










Serviços Personalizados
Journal
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de fitopatología
versão On-line ISSN 2007-8080versão impressa ISSN 0185-3309
Rev. mex. fitopatol vol.33 no.2 Texcoco 2015
Scientific Articles
Anthracnose ( Colletotrichum gloeosporioides Penz.) of litchi fruit ( Litchi chinensis Soon.) in Oaxaca, México
1Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Campo experimental Rosario Izapa. Km. 18 carretera Tapachula-Cacahoatán, Tuxtla chico, Chiapas; México.
2Colegio de Postgraduados, km. 36.5 carretera México-Texcoco, Montecillo, Texcoco, CP 56230, México.
3Colegio de Postgraduados, km. 88.5 carretera Xalapa-Veracruz, CP 91690 Tepetates, Manlio Fabio Altamirano, Mexico.
4Universidad del Papaloapan. Campus Tuxtepec. Circuito Central # 200. Colonia Parque Industrial. CP 68301, Tuxtepec, México.
Anthracnose of litchi fruits was characterized by internal and external brown circular lesions with well-defined margins on the pericarp, but not on the aril or seed. Colletotrichum sp., Pestalotiopsis sp., Fusarium sp., and Lasiodiplodia sp. were isolated from symptomatic fruit pericarp samples of cultivars Mauritius and Brewster in 2008-2010 cycles. Colletotrichum sp. (51 %) and Pestalotiopsis sp. (17 %) were isolated more frequently from symptomatic fruit pericarp samples of cultivar Mauritius in 2009. Colletotrichum sp. was isolated on PDA (39 %) and in semi-selective medium (59 %) from symptomatic Mauritius fruit pericarp samples in 2010, and from asymptomatic pericarp Brewster samples in 2009 (32 %) and 2010 (21 %) on PDA and 10 % in semi-selective medium; however it was not isolated from asymptomatic Mauritius pericarp in neither of the two harvesting years. Only Colletotrichum sp., reproduced the disease in the laboratory (17 out of 20 inoculated fruits) and in the field (19 out of 20 inoculated fruits). The fungus was morphologically and molecularly identified as Colletotrichum gloeosporioides Penz.
Keywords: Brewster; incidence; Mauritius; pathogenicity; PCR; pericarp
La antracnosis de frutos de litchi se caracterizó como lesiones internas y externas, obscuras, circulares con márgenes definidos en el pericarpio, pero no en el arilo y semilla. Colletotrichum sp., Pestalotiopsis sp., Fusarium sp., y Lasiodiplodia sp., se aislaron de muestras de pericarpio sintomático de los cultivares Mauritius y Brewster en los ciclos 2008-2010. Colletotrichum sp. (51 %) y Pestalotiopsis sp. (17 %) se aislaron más frecuentemente de muestras de pericarpio de frutos sintomáticos del cultivar Mauritius en 2009. Colletotrichum sp., se aisló en PDA (39 %) y en medio semiselectivo (59 %) de muestras de pericarpio sintomático en el cultivar Mauritius en 2010, y de pericarpio asintomático del cultivar en 2009 (32 %) y 2010 (21 %) sobre PDA y 10% en medio semiselectivo; sin embargo no se aisló de pericarpio asintomático del cultivar Mauritius en ninguno de los dos años de cosecha. Sólo Colletotrichum sp., reprodujo la enfermedad en laboratorio (17 de 20 frutos inoculados) y en campo (19 de 20 frutos inoculados). El hongo se identificó morfológica y molecularmente como Colletotrichum gloeosporioides Penz.
Palabras clave: Brewster; incidencia; Mauritius; patogenicidad; PCR; pericarpio
Palabras clave: Brewster; incidencia; Mauritius; patogenicidad; PCR; pericarpio
Introduction
Litchi (Litchi chinensis Soon) is a subtropical fruit (Coates et al., 1994) native to southern China and southeast Asia; its pericarp is thin and rough, and when fruits get ripe, it becomes red and cracked due to the presence of brachysclereids, which could be associated with protection against mechanical damage or physiological stress (Nacif et al., 2001). The aril is white and covers a dark brown seed.
The main problems in litchi production are low or irregular yields due to excessive fruit drop (Mitra et al., 2005); oxidation and cracking of the pericarp (Huang et al., 2001; Xu et al., 2005); pest damages caused by C. gloeosporioides Penz (Campbell and Campbell, 2001), Peronophythora litchii Chen Ex Ko, Tessaratoma papillosa Drury and Dasineura sp., and high harvesting and packaging costs (Xu et al., 2005).
Litchi production in Mexico is 19,888.05 tons, in an area of 3,738.48 hectares (SIAP, 2013). In Oaxaca, Mexico, Brewster and Mauritius cultivars are commercially grown in the Papaloapan and Bajos Mixes regions, respectively. In both cultivars, a regular dark brown spot on the pericarp represents one of the main problems under field and postharvest conditions. Field symptoms are characterized by brown and dry semicircular lesions; the aril and seed show no damages. No records have been found about its cause and importance in Mexico so far; however, Oaxaca farmers report damages of up to 50% in production. Taking in to account the socioeconomic importance of this problem, the objective of this study was to determine the etiology of the brown spot of litchi fruits in Oaxaca, Mexico.
Materials and Methods
The experiment was established in commercial litchi orchards in three counties of Oaxaca State during the 2008-2010 production cycles. Brewster and Mauritius cultivars were evaluated in San Jose Chiltepec (17° 57' 57.92" N, 96° 10' 11.68" W), and in San Juan Cotzocón (17° 26' 07.74" N, 95° 19' 53.97" W) and Loma Bonita (17° 58' 53.1" N, 95° 50' 58.1" W), respectively.
Symptoms and fungi characterization. Symptomatic (natural infection) and asymptomatic commercial ripe fruits from Brewster and Mauritius cultivars were collected in May 2009 in one commercial litchi orchard in San José Chiltepec and San Juan Cotzocón, respectively. One hundred fruits from each cultivar were randomly collected from 25 trees and then, washed with soap and water in the laboratory, air dried at room temperature, and longitudinally sectioned. Symptom consistence, shape, and color were evaluated on pericarp, aril, seed, and peduncle, as well as the presence of chlorotic haloes and signs of associated microorganisms.
To characterize fungal isolates, 123 symptomatic and 195 asymptomatic Brewster fruits were harvested in 2008-2009, and 60 and 70 in 2009-2010. Then, 193 symptomatic and 72 asymptomatic Mauritius fruits were sampled in 2008-2009, and 102 and 96 in 2009-2010. Two 1.5 cm2 sections of pericarp and aril were dissected from each fruit and disinfested in 1.5 % sodium hypochlorite for three min, rinsed three times in distilled water for 1 min each, and dried with sterile paper towels. Tissue sections were cultured on PDA (Bioxon(tm)) and on a Colletotrichum semi-selective medium [19.5 g PDA, 7.5 g agar (Merck(tm)), 8 mg iprodione, 20 mg triadimefon, 50 mg chloramphenicol (Merck(tm)), and 200 mg streptomycin ] (Davis, 2003) for 5-7 days at 25 °C under fluorescent light. Fungal growths were purified by hyphal tip and monoconidial cultures.
Pathogenicity tests. Colletotrichum, Pestalotiopsis, Fusarium, and Lasiodiplodia were isolated from all the Brewster and Mauritius pericarp symptomatic samples and their pathogenicity was tested under laboratory and field conditions.
Laboratory pathogenicity test was evaluated in asymptomatic and commercially ripe Mauritius fruit collected in field in May 2010. Twenty disinfested fruits were wounded with a sterile needle and a 0.5 cm agar disc with Colletotrichum, Pestalotiopsis, Fusarium, or Lasiodiplodia was placed on each wound (Johnson and Cooke, 2002). Control fruits (mockinoculated) were inoculated with a 0.5 cm agar disc without mycelia. All inoculated fruits were maintained under humidity chamber conditions at 28 °C±2 for 7 days. Observations were done every 24 h to detect symptoms.
Pathogenicity under field conditions was evaluated in May and June 2010 on 20 asymptomatic Mauritius fruits per tree (5 fruits/cardinal point). Two development stages were evaluated: at the beginning of ripening, and commercially ripe fruits. Fruits were disinfested with 70 % ethanol and wounded with a sterile needle in the middle of the fruit and at the base of the peduncle. Fruits were individually inoculated at evening (18:00 - 20:00 central time) by injecting 0.5 ml of a 1 x 106 conidia.ml-1 suspension of Colletotrichum, Pestalotiopsis, Fusarium, Lasiodiplodia or sterile distilled water (mockinoculated) (Dinh et al., 2003). Inoculated wounds were covered with wet sterile cotton for 24 h to create high humidity conditions for fungal growth. Symptom development was registered for 15 days. Re-isolates from laboratory and field infected fruits were made to verify Koch's postulates.
Morphological characterization of the pathogen. Four out of 256 Colletotrichum isolates obtained from symptomatic fruits were selected based on their mycelia growth and spore production, and designed as M26, M28, M29, and M30. Genus (Sutton, 1980; Hanlin, 1990; Barnett and Hunter, 2006) and species (Sutton, 1980; Smith and Black, 1990; Photita et al., 2005) were determined from monoconidial colonies. Appressorium formation was induced on PDA cover slides inoculated individually with M26 and M28-M30 spores (Johnston and Jones, 1997). Appressorium shape and size were characterized a 7 day old fungal culture on PDA.
M26 and M28-M30 growth rates were evaluated for seven days on PDA plates cultured at 28 °C. Data were statistically analyzed by ANOVA (α=0.05) and grouped by Tukey test (SAS, 1997).
Molecular identification. M26 and M28-M30 DNA was isolated following the AP method (Sambrook and Russel, 2001). DNA integrity was electrophoresed on a 1% agar gel at 80 volts for 40 min in TBE buffer, and stained with ethidium bromide. ITS1 and ITS2 regions were amplified by PCR with primers ITS4 (5′-TCCTCCGCTTATTGATATGC-3′) and ITS5 (5′-GGAAGTAAAAGTCGTAACAAGG-3′) (White et al., 1990), which amplify a 580 bp fragment. To identify and confirm species, DNA was amplified with ITS4 and the CgInt (5′-GGCCTCCCGCCTCCGGGCGG-3′) specific primer (Mills et al., 1992), which amplify a 450 bp fragment.
PCR amplification was done in a thermocycler (Applied BiosystemsTM 2720) in a 25 µl reaction with 1.0 µl of each primer (10 mM), 1.0 µl of dNTP (10 mM), 5 µl of PCR Buffer 10X, 2.5 µl of MgCl2 (50 mM), 0.5 µl of Taq Polymerase, 0.5 µl of DNA and 13.5 µl of nuclease-free water. PCR reactions without tempered were used as negative control.
PCR consisted of an initial denaturation of 5 min at 95 °C followed by 35 cycles of 1 min of denaturation at 95 °C, 1 min of annealing at 57 °C, and 1 min of extension at 72 °C, with a final extension of 10 min at 72 °C. PCR products were electrophoresed on a 1 % agarose gel, purified with the QUIAGEN DNeasy Plant Kit and sequenced in a 3730XL automated sequencer. The sequences were lined up against sequences in the gene bank and homology was determined in the National Center for Biological Information (BLAST, 2011).
Results
Symptom characterization. The Brewster exocarp showed a dark brown lesion without chlorotic halo (Figure 1A). On the endocarp, the lesion was a dark brown circular, and semi-humid depressed spot without chlorotic halo (Figure 1B). On Mauritius exocarp it developed a dark brown well defined lesion with chlorotic halo (Figure 1D); and on the endocarp a dry dark brown non-depressed spot was identified (Figure 1E). None of the Brewster and Mauritius fruits showed damages on the aril (Figure 1C and 1F ), seed, or peduncle, and no signs of microorganisms were observed.

Figure 1 Litchi fruits (Litchi chinensis Soon) with anthracnose symptoms. Brewster cv: A) Pericarp external symptom, B) Asymptomatic aril, C) Pericarp internal symptom. Mauritius cv: D) Pericarp external symptom, E) Asymptomatic aril F) Pericarp internal symptom. Pathogenicity of C. gloeosporioides on Mauritius litchi fruits in the laboratory: G) Control fruits, H) Pericarp external symptom, I) Pericarp internal symptom. Pathogenicity on Mauritius litchi fruits in the field: J) Control fruits, K) Pericarp external symptom, L) Pericarp internal symptom. Bar = 1 cm.
Fungi isolation. Colletotrichum sp., Pestalotiopsis sp., Fusarium sp., and Lasiodiplodia sp. were isolated from symptomatic Brewster and Mauritius pericarps in the 2008-2009 and 2009-2010 production cycles. However, Colletotrichum sp. (256/640 isolates) and Pestalotiopsis sp. (71/640 isolates) were isolated more frequently than the other ones (Table 1 and 2 ). Colletotrichum sp., Pestalotiopsis sp., and Lasiodiplodia sp. were also isolated from Brewster asymptomatic pericarps (Table 1); and Fusarium from asymptomatic Mauritius samples (Table 2). Colletotrichum sp. grew on more pericarp tissue fragments incubated on semi-selective medium than those cultured on PDA (Table 1 and 2 ). Colletotrichum sp. isolates produced conidia on 4 and on 8-12 day old semi-selective and PDA cultures, respectively. No microorganisms were isolated from aril tissue fragments incubated on both growth media (Table 1 and 2 ).
Table 1 Fungal genera isolates from symptomatic and asymptomatic litchi fruits (Brewster cultivar), in San Jose Chiltepec, Oaxaca.
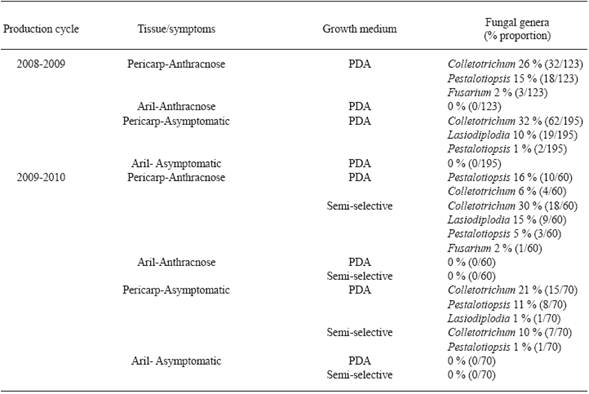
Table 2 Fungal genera isolates from symptomatic and asymptomatic litchi fruits (Mauritius cultivar), in San Juan Cotzocón, Oaxaca.
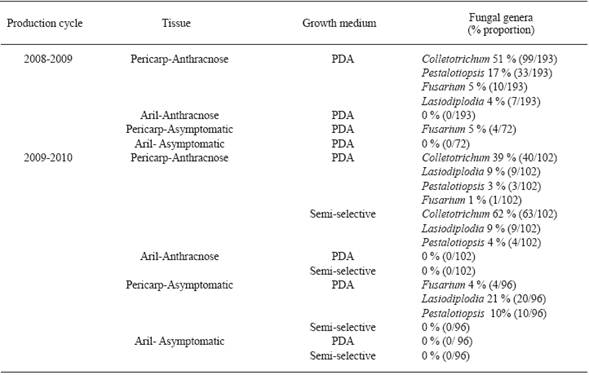
In 13 of 15 sections of pericarp incubated in humidity chamber there was conidia production on the fifth day; the salmon colored conidia masses were morphologically identified as Colletotrichum sp.
Pathogenicity tests. Laboratory test results showed that 85 % (17 out of 20) of the fruits inoculated with Colletotrichum sp. developed symptoms on the pericarp similar to those observed in the field. These symptoms appeared 72 h after inoculation as a small light brown circular lesion (Figure 1G). On the endocarp it was developed a dark brown circular and semi-humid lesion (Figure 1H). Mock inoculated fruits (Figure 1I) and fruits inoculated with Fusarium sp., Pestalotiopsis sp., or Lasiodiplodia sp. did not show any symptom.
In the field, symptoms developed in 95 % (19 out of 20) of the commercial ripe Mauritius fruits inoculated with Colletotrichum sp. on the middle of the pericarp in May, and 90 % in June. Symptoms were observed eight days after inoculation as dark brown lesions on the pericarp (Figure 1J). Symptom on the endocarp was a dark brown circular and depressed lesion (Figure 1K).
Mock inoculated fruits (Figure 1L) and fruits inoculated with Fusarium sp., Pestalotiopsis sp., or Lasiodiplodia sp. did not show any symptoms.
The Colletotrichum isolate that induced anthracnose symptoms under laboratory and field conditions was re-isolated and identified morphologically as C. gloeosporioides (Sutton, 1980).
Pathogen characterization and identification. Most (193) of the 256 isolates of C. gloeosporioides formed white-light gray scarcely cottony mycelia with abundant bright orange conidial masses in concentric circles (Figure 2A, 2D and 2G ). Some (78) of them produced white mycelium with very few conidial masses near to the transference point (Figure 2J). Four out of these 256 isolates were designed as M26, M28, M29 and M30.
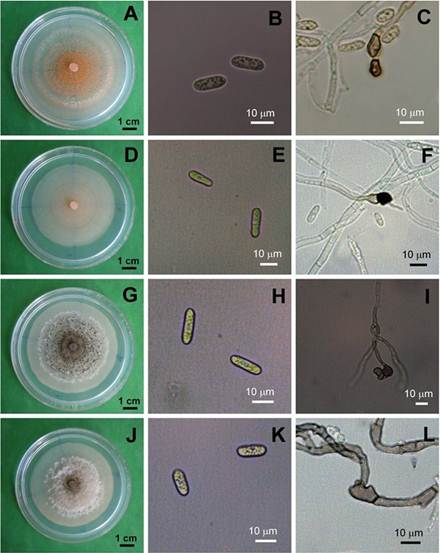
Figure 2 Colony Morphology (A,D,G, J), conidia (B, E H, K) and mycelial appressoria (C, F, I, L) of the four representative isolates M26 (A,B,C), 28 (D,E,F), 29 (G,H,I), 30 (J,K.L) of Colletotrichum gloeosporioides from litchi fruits, after five days in PDA.
M26 and M28-M30 conidia were cylindrical (Table 3 and Figure 2B, 2E , 2H and 2K ). M26 conidia were statistically longer than M28 ones, which had the greatest diameter (Table 3). The four isolates formed clavate appressoria (Figure 2C, 2F , 2I and 2L ). There were not differences among the mycelium growth rates of the four isolates cultured at 28 °C on PDA (α= 0.05) (Table 3).
Table 3 Conidium size and mycelium growth rate of M26 and M28-M30 C. gloeosporioides isolates obtained from litchi fruits with anthracnose symptoms in San Jose Chiltepec and Loma Bonita, Oaxaca (2009-2010 production cycle).
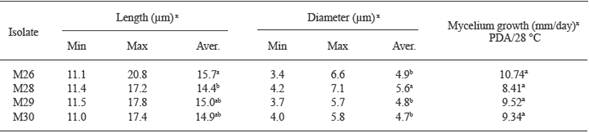
x Values with the same letter in each row are statistically the same, Tukey (P≤0.05).
Molecular identification. ITS4-ITS5 and ITS4-CgInt primers amplified a fragment of 580 and 450 bp, respectively, which indicated that M26 and M28-M30 isolates belong to C. gloeosporioides (Figure 3).
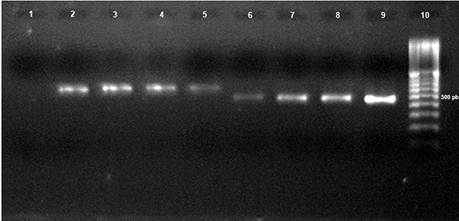
Figure 3 PCR product of four characteristic isolates (M26, M28, M29, and M30) of Colletotrichum gloeosporioides. Lines: 1) Control (H2O PCR); 2) M26- ITS4/ITS5; 3) M28- ITS4/ITS5; 4) M29- ITS4/ITS5; 5) M30- ITS4/ITS5; 6) M26- ITS4-CgInt; 7) M28- ITS4-CgInt; 8) M29- ITS4-CgInt; 9) M30- ITS4-CgInt; 10) 100 base pair (bp) marker (Invitrogen).
M26 and M28-M30 amplicons were sequenced and aligned with those in the GenBank, and gave a homology of 96, 93, 99, and 99 % with C. gloeosporioides, respectively (gb/HM016794.1; gb/JF487788.1; gb/JF487788.1; gb/FJ755268.1) (BLAST, 2011).
Discussion
The results of this study indicate that C. gloeosporioides is the causal agent of anthracnose disease in pre-harvest litchi fruits of Brewster and Mauritius cultivars in Oaxaca, Mexico. The fungus was isolated from natural and artificially infected fruits of both cultivars, and formed a cottony white to light gray mycelial growth with salmon colored conidial masses on PDA. Symptoms on inoculated fruits were very similar to those observed in field with natural infection.
In this research, Colletotrichum was also isolated from asymptomatic Brewster fruits; similar results were reported by Ann et al., (2004), who isolated C. gloeosporioides from asymptomatic flowers and immature fruits of the Black leaf litchi cultivar. Once fruits were harvested and stored, they developed anthracnose symptoms, suggesting that this pathogen attacks litchi through latent infections. We consider the probable endophytic nature of C. gloeosporioides in the field.
As mentioned before, litchi anthracnose has been consigned mainly as a postharvest disease (Coates et al., 1994, Liu et al., 2003, and Liu et al., 2006) that results from quiescent infections (Li et al., 2005); however, this fungus has been reported as the causal agent of the peduncle rot and pepper spotting on litchi fruits under field conditions in Australia (Cooke et al., 2009). Others causes of pericarp browning at postharvest are moisture loss due to desiccation of the pericarp (Chen and Hong, 1992; Underhill and Critchley, 1993), degradation of anthocyanins and oxidation of phenolic substrates by polyphenol oxydase (Underhill and Critchley, 1993) and/or peroxidase enzymes (Zhang and Quantick, 1997).
Species of Colletotrichum cannot be recognized by host range (Sutton, 1980, 1992), so identification and characterization of the isolates obtained in this study were done based on morphological characteristics (Smith and Black, 1990; Gunnell and Gubler, 1992; Sutton, 1992). According to those criteria four out of 256 isolates of Colletotrichum spp. were selected (M26, M28-M30) and identified as C. gloeosporioides Penz; their micelial growth pattern agree with that described by Alvarez (1949); however, according to Smith and Black (1990) and Gunnel and Gluber (1992), this growth is similar to C. fragariae grown on PDA, but C. fragariae cultures on cornmeal, oatmeal, or bean-lime agar plates are dark with concentric circles and pink colored conidial masses (Brooks, 1931). In adition, the average size of the M26, and M28-M30 conidia were shorter and wider than those reported in C. fragariae (Smith and Black, 1990; Gunnel and Gubler, 1992).
Based on PCR amplifications through specific species primers, M26 and M28-M30 species were corroborated as C. gloeosporioides with a 95-100% similarity with sequences reported for the same species in the gene bank (Freeman et al., 2000).
Conclusions
Anthracnose symptoms observed in litchi fruits in Oaxaca were caused by Colletotrichum gloeosporioides Penz. The disease was characterized by dark-brown and circular dry lesions on the exocarp and endocarp, but neither in the aril nor seed. This is the first report of Colletotrichum gloeosporioides as the causal agent of anthracnose of litchi fruits (Litchi chinensis) in Oaxaca, México.
REFERENCES
Alvarez GLA. 1949. Anthracnose of the Annonaceae in Puerto Rico. Journal of Agriculture of the University of Puerto Rico 33: 27-43. [ Links ]
Ann PJ, Tsai JN, Wang IT and Young HR. 2004. Litchi fruit anthracnose and the cause of black skin. Plant Pathology Bulletin 13: 299-308. [ Links ]
Barnett LH and Hunter BB. 2006. Illustrated Genera of Imperfect Fungi. Minessota. The American Phytopathological Society Press. 218 p. [ Links ]
BLAST(r). Basic Local Alignment Search Tool. Available at: Available at: http://blast.ncbi.nlm.nih.gov/Blast.cgi . Accessed on July 13, 2011. [ Links ]
Brooks AN. 1931. Anthracnose of strawberry caused by Colletotrichum fragariae, n. sp. Phytopathology 21: 739-744. [ Links ]
Campbell RJ and Campbell CW. 2001. Evaluation and introduction of lychee cultivars in South Florida, USA. Acta Horticulturae 558: 119-123. [ Links ]
Chen HJ and Hong QZ. 1992. A study on the senescence and browning in the pericarp of litchi (Litchi chinensis Sonn) during storage. Acta Horticulturae Sinica 19: 227-232. [ Links ]
Coates LM, Johnson GI, Sardsud U and Cooke AW. 1994. Postharvest diseases of lychee in Australia, and their control. In: Johnson GL and Highley E (Eds.) Development of Postharvest Handling Technology for Tropical Tree Fruits: A Workshop Held. Bangkok: ACIAR. pp. 68-69. [ Links ]
Cooke T, Persley D and House S. 2009. Diseases of Fruit Crops in Australia. Queesland. CSIRO Publishing. Pp: 145-148. [ Links ]
Davis MJ. 2003. Lychee anthracnose inoculums sources. Proceedings of the Florida State Horticultural Society 116: 12-15. [ Links ]
Dinh SQ, Chongwungse J, Pongam P and Sangchote S. 2003. Fruit infection by Colletotrichum gloeosporioides and anthracnose resistance of some mango cultivars in Thailand. Australasian Plant Pathology 32: 533-538. [ Links ]
Freeman S, Minz D, Jurkevitch E, Maymon M and Shabi E. 2000. Molecular analyses of Colletotrichum species from almond and other fruits. Phytopathology 90: 608-614. [ Links ]
Gunnell PS and Gubler WD. 1992. Taxonomy and morphology of Colletotrichum species pathogenic on strawberry. Mycologia 84: 157-165. [ Links ]
Hanlin RT. 1990. Illustrated Genera of Ascomycetes. Minnesota. American Phytopathological Society Press. 263 p. [ Links ]
Huang X, Li J, Wang H, Huang H and Gao F. 2001. The relationship between fruit cracking and calcium in litchi pericarp. Acta Horticulturae 558: 209-215. [ Links ]
Johnson GI and Cooke AW. 2002. Postharvest disease control in Lychee. Acta Horticulturae 575: 705-715. [ Links ]
Johnston PR and Jones D. 1997. Relationship among Colletotrichum isolates from fruit rots assessed using rDNA sequences. Mycologia 89: 420-430. [ Links ]
Li X, Liu A and Chen W. 2005. Studies on development and control of anthracnose of lychee fruit before and after harvest. Acta Horticulturae 665: 409-413. [ Links ]
Liu A, Chen W and Li X. 2006. Developments of anthracnose on harvested litchi fruits and the effects of the disease on storage of the fruits. Acta Phytopathologica Sinica 33: 351-356. [ Links ]
Liu A, Chen W and Li XY. 2003. Biological characteristics of the pathogenic fungus causing anthracnose of litchi. Acta Phytopathologica Sinica 33: 313-316. [ Links ]
Mills PR, Sreenivasaprasad S and Brown AE. 1992. Detection and differentiation of Colletotrichum gloeosporioides isolates using PCR. FEMS Microbiology Letters 98: 137-144. [ Links ]
Mitra SK, Pereira LS, Pathak PK and Majumdar D. 2005. Fruit abscission pattern on lychee cultivars. Acta Horticulturae 665: 215-218. [ Links ]
Nacif SR, Paoli AAS and Salomão LCC. 2001. Morphological and anatomical development of the litchi fruit (Litchi chinensis Sonn. cv. Brewster). Fruits 56: 225-233. [ Links ]
Photita W, Taylor PWJ, Ford R, Hyde KD and Lumyong S. 2005. Morphological and molecular characterization of Colletotrichum species from herbaceous plants in Thailand. Fungal Diversity 18: 117-133. [ Links ]
Sambrook J and Russel DW. 2001. Molecular Cloning. A Laboratory Manual Third edition. New York. Cold Spring Harbour Laboratory Press. 46 p. [ Links ]
SAS Institute. 1997. The SAS System for Windows. Release 6.12. Cary, NC, USA: SAS Institute Inc. [ Links ]
SIAP. Servicio de Información Agroalimentaria y Pesquera. Available at: Available at: http://www.siap.gob.mx/agricultura-producción-anual/ . Accessed on June 29, 2015. [ Links ]
Smith BJ and Black LL. 1990. Morphological, cultural and pathogenic variation among Colletotrichum species isolated from strawberry. Plant Disease 74: 69-76. [ Links ]
Sutton BC. 1980. The Coelomycetes: Fungi Imperfecti With Pycnidia Acervuli and Stroma Surrey. Cabi Publishing. 696 p. [ Links ]
Sutton BC. 1992. The genus Glomerella and its anamorph Colletotrichum. In: Bailey JA and Jeger MJ (Eds.) Colletotrichum: Biology, Pathology and Control. Wallingford. CAB International. pp: 1-26 [ Links ]
Underhill SJR and Critchley C. 1993. Physiological, biochemical and anatomical changes in lychee (Litchi chinensis Sonn.) pericarp during storage. The Journal of Horticultural Science and Biotechnology 68: 327-335. [ Links ]
White TJ, Bruns T, Lee S and Taylor J. 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JH and White TJ (Eds.) PCR Protocols. Academic Press. Pp: 315-322. [ Links ]
Xu X, Wu Z, Chen W and Su M. 2005. Effect of low temperature on relative enzyme activity and membrane peroxidization of lychee fruits. Acta Horticulturae 665: 359-364. [ Links ]
Zhang DL and Quantick PC. 1997. Effects of chitosan coating on enzymatic browning and decay during postharvest storage of litchi (Litchi chinensis Sonn.) fruit. Postharvest Biology and Technology 12: 195-02. [ Links ]
Received: November 20, 2014; Accepted: June 28, 2015
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
