










Serviços Personalizados
Journal
Artigo
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkGaceta médica de México
versão On-line ISSN 2696-1288versão impressa ISSN 0016-3813
Gac. Méd. Méx vol.140 no.2 Ciudad de México Mar./Abr. 2004
Actividades académicas
DNA Medio Siglo.
II. Preámbulo histórico del DNA
Roberto Kretschmer*
*Dr.Roberto Kretschmer S. FAAAP, FAAAAI. Director de la Unidad de Investigación Médica e Inmulogía, Centro Médico Nacional, Siglo XXI, Instituto Mexicano del Seguro Social. México, D.F.
La teatral espectacularidad de la estructura de hélice doble del DNA propuesta por Watson y Crick en 1953, modelo capaz de almacenar y -replicándose- de transmitir información genética, podría hacer pensar que llegó como Epifanía. Nada más falso. Sobre hombros de gigantes y siguiendo un largo camino que a veces parecería una Odisea, Watson y Crick pisaron terreno muy sólido cuando lo construyeron usando datos ajenos. Veamos.
El origen de las Especies de Darwin (1853) avivó la curiosidad por aclarar los mecanismos de la herencia, y dejó atrás al simpático homúnculo. Gregor Mendel, monje agustino -como Lutero- literalmente se tropieza con la herencia dominante y recesiva hibridizando chícharos en la hortaliza del Convento de Nuestra Señora en Brünn/Brno (Moravia) y lamentando que sus anhelados híbridos no resultaran definitivos. Al mismo tiempo el joven Friedrich Miescher (Figura 1) de la estirpe biomédica de los Miescher y de los His de Basilea va a Tübingen, a orillas del Neckar, para hacer bioquímica, la nueva ciencia, la ciencia del futuro, con el famoso Felix Hoppe-Seyler. En el austero laboratorio (Figura 1) de la fortaleza (de hecho había sido la cocina del castillo), nunca antes rozada por la historia (excepto que en sus laderas languideciera por 40 años en su locura el poeta Hölderlin), debe indagar lo que contienen los núcleos de las células. Miescher los extrae de los leucocitos de los apositos llenos de pus obtenidos de la Clínica Quirúrgica -aún no existían ni antisepsia ni antibióticos- y responde:
<<....... este material viscoso ...... no es ni proteína, ni azúcar, ni grasa (los pilares químicos de la materia viva) aunque tiene un poco de todos ellos, sobre todo proteína...>> , que denomina protamina. Rica en fosfato, la cree el almacén de fósforo en el cuerpo, y por tornarse amarilla con ácido nítrico, concluye que poseía xantinas, luego tan estudiadas por Emil Fischer. La bautizó nucleína. Cauto, como buen profesor alemán, Hoppe-Seyler procrastinó dos años, pero al fin cede, y el trabajo aparece en 1871. Esa nucleína devendrá en el DNA. Al final de su vida Miescher intuyó que la nucleína podría ser el substrato químico de la herencia. Lo publicado por Mendel en 1866 en las Abhandlungen der Naturwissenschafilichen Gesellschaft zu Brünn , revista seguramente no leída por muchos (ahora diríamos que con un impacto bajísimo), tuvo que ser rehabilitado por De Vries, Correns y von Tschermack 30 años más adelante, para ya quedarse como leyes, y el mismo Mendel como el santo patrono secular de la genética moderna. Miescher por otra parte tendría un seguimiento más inmediato, gracias a Albrecht Kossel (Figura 2), otro alumno de Hoppe-Seyler, activo en Estrasburgo y Heidelberg y premio Nobel en 1910. Obsesivo, gran químico y pacifista, Kossel describe los cuatro ácidos nucléicos (adenina, guanina, citosina y timina) presentes en la nucleína, y pronostica la pentosa -azúcar luego confirmado por Hammersten. Pero la molécula integrada (i.e. ácido desoxirribonucléico = DNA ) aún no la concibe. Descubre sin embargo lo que ahora es obvio: que la nucleína era lo mismo, no importa su origen (i.e. humanos, insectos, peces, vegetales, bacterias, etcétera) .....i..... como son ........ lo mismo ......todos los libros de una biblioteca!


La primera mitad del siglo XX, vena grandes avances en la bioquímica de la nucleína, sobresaliendo en ello, Febo Aarón Levene (Figura 3), quien estudiara con Pavlov y con el músico-quimico Borodín en Rusia, y luego con el ya citado Kossel, antes de emigrar a América. Él, y más tarde Todd, desmenuzan químicamente al DNA: enlaces, distancias interatómicas, ángulos planares, cargas, etcétera. Irónicamente, la molécula de DNA más lograda por estos químicos, resultó ser un tetranucleótido: cerrado, inmutable ......perfecto (Figura 3). Pero demasiado pequeño como para tener que ver algo con la herencia, que -se creería cada vez más- sólo podría recaer en moléculas mucho más grandes, como las proteínas. El pobre Febo cargará por ello con la culpa histórica de haber propuesto una molécula tan incuestionable (el tetranucleótido), que así se convierte en barrera, que no camino, en la búsqueda de la estereoquímica macromolecular del DNA. Los genes (o como se les llamara entonces: gémulas, idioplasmas, parigenes, bióforos, etc. segaran siendo una entelequia. En esta fase histórica del DNA, la última aportación de la Bioquímica (regresará eso si, casada con la Genética, ya como Biología Molecular) será la notable observación de Erwin Chargaff (Figura 4) de que en las células las concentraciones molares de adenina siempre son iguales a las de timina, y las de citosina iguales a las de guanina. Ésta, la llamada Regla de Chargaff fue la primera insinuación -que no certeza- de complementariedad, y le resultará crucial a Watson y Crick al proponer la función codificadora del DNA. En los años cuarenta Caspersson revive el estudio del DNA al señalar que no se trataba de una molécula pequeña, sino de un polimero enorme y birrefringente. Más grande -pero aparentemente también menos complejo- que las proteínas. Ahora sabemos porqué: DNA = cuatro bases, fosfato y deoxirribosa, en tanto que proteínas: mezcla de 20 posibles aminoácidos y algunos agregados (azúcar, lípidos).


Todo esto ocurre mientras la genética se va insinuando en la literatura: los casos de inexorable alcoholismo en Zola, y la curiosa persistencia de buena dicción de un paria aristocrático en Dickens, como inescapables dictados de la herencia. Tristemente, la genética desciende también a los abismos humanos con las estupideces de Lysenko en la URSS, la criminal eugenia de los nazis y algún pecadillo americano en Ellis Island.
En los años veinte, Morgan (Premio Nobel en 1933) con sus modestas y libidinosas moscas ( Drosofila melanogaster ), de tan sólo cuatro cromosomas, demostrará que éstos sí albergan a los genes. Los cautos Avery, Macleod y McCarty en los años cuarentas, y usando los aún más modestos neumococos, aclaran que los genes (y por ende los cromosomas) contienen DNA, y es el DNA -y no una proteína- el material de la transformación genética. Sorprendentemente el premio Nobel los esquiva.
Aquí, hay que hacer un descanso en el camino, para referirnos a un evento extraordinario, casi milagroso en la historia de la ciencia. Una transferencia epistemológica inédita, que llevará directamente a la inteligente macromolécula que es el DNA. Algunos físicos teóricos ya galardonados con el Nobel, la créme de la créme de la Ciencia, mostraron en los años treintas y cuarentas un creciente interés por los fenómenos biológicos. Habiendo explicado ya la Materia y la Energía, aplican ahora la mecánica cuántica a la Vida. El origen parece haber sido la obscura conferencia " Luz y Vida " que dictara el gran físico danés Niels Bohr (Premio Nobel 1922) en 1932, y cuyo mensaje recoge Max Delbrück (Nobel con Luria en 1969) (Figura 5), un físico alemán quien al regresar a Berlín desarrolla un modelo cuántico de mutación genética en moscas. Se marcha a California, pero como las moscas de Morgan le parecen enormes, opta por formas de vida más simples y en cierta forma aún más libidinosas: las bacterias. Con Salvatore Luria (Figura 5) un físico italiano cercano a Fermi, el mercurial físico húngaro Szillard, y otros forman el Grupo de los Fagos y confirman elegantemente lo propuesto por Avery: sólo el DNA de los fagos -i.e. virus que infectan bacterias- penetra en las bacterias y las muta. La proteína del fago se queda invariablemente afuera (q. e. d!).

Erwin Schrödinger (Premio Nobel 1933) (Figura 6), físico austriaco, emigra a la católica Irlanda y en 1944 publica su notorio librito "What is Life?" (Figura 6), mientras en su casa en Dublín convive pacíficamente con su esposa en el primer piso y su amante en la planta baja "i....... now...... that is life! " .......! Humor aparte, este pequeño libro causó un gran -aunque controversial- impacto en los científicos de la época. Schrödinger señala que los genes podrían ser cristales aperiódicos en un cubo de 3 Å por lado y 1000 átomos por gene; que funcionaría como un código Morse binario, con instrucciones para la vida, y susceptible de sufrir mutaciones mínimas (i.e. cuánticas) y estables, menú de propuestas para la selección natural ........el lenguaje de la evolución.

Ciertamente, a final de cuentas el DNA resultó ser un cristaloide aperiódico, un código mas bien terciario (i.e. codones, vide infra ) a base de tres de cuatro símbolos (A, G, C, T, vide ante ). iComo podemos ver, Schrödinger no nada más era hábil con las chicas! Más de un físico se hizo biólogo por sus ideas.
Sin embargo, lo más trascendental de esta notable transferencia epistemológica, lo fueron los difractores de rayos X. Max von Laue físico alemán (Premio Nobel 1914), demuestra en 1906 que en su paso por un cristal, los rayos X se desvían sólo por los caminos que les permiten los átomos de la molécula, y estudia así las propiedades de los rayos X. Los Bragg, físicos ingleses -y único caso de premio Nobel (1915) compartido por padre e hijo- hacen lo opuesto: ahora estudian la estructura de un cristal en base al patrón de difracción que provoca en los rayos X.
Final de la Segunda Guerra Mundial: Randall, inventor del radar y por ello héroe de la batalla de Inglaterra, usa su influencia para promover visionariamente la difracción de los rayos X pero ahora aplicada a moléculas de interés biológico, como se lo sugiriera años antes John Bemal, el multifacético irlandés hispano-sefaradí: Proteínas (hemoglobina y mioglobina) en el Instituto Cavendish de Cambridge (Perutz, Kendall y Crick), y DNA en King's College de Londres (Rosalind Franklin y Wilkins). Incidentalmente casi todos ellos físicos, y todos -excepto Rosalind Franklin, por su muerte prematura (1958), y no por chauvinismo- recibirán un premio Nobel en 1962.
En Califomia, Linus Pauling (Figura 7) posiblemente el físico químico más notable del siglo XX crea la escuela americana de difracción de rayos X y describe las α-hélices, una estrategia estructural usada por muchas proteínas. Pero sobre todo, y esa es la personalísima aportación de Pauling, el juego -como un Tinkertoy® intelectual- de construir modelos moleculares tridimensionales plausibles, con estricto apego a las reglas que imponen los datos químicos y el patrón de difracción de los rayos X por la sustancia.

Con empujones de Sziflard y Luria llega al Instituto Cavendish, desertor de Copenhague, el biólogo otrora ornitólogo, James Watson, un joven de 23 años un tanto impulsivo, desaliñado e irreverente, típico ejemplo de midwestener americano de cepa irlandesa. De la escuela de los fagos, inmediatamente hace " click " con el efusivo Crick, 12 años mayor que él y empiezan a jugar -como Pauling- a los modelos físico-químicos del DNA, cuando no están jugando tenis o remando en el río Cam. De paso generan una gran indignación institucional, enfática por parte de Lawrence Bragg y de Franklin. Ésta llegó a exigir que se pusiera fin a esos juegos pedestres -que además no les correspondían temáticamente- hasta no tener datos más definitivos, y propios. En el austero instituto, se llegó inclusive a bautizar sarcásticamente el cuarto de Watson y Crick como ........"el WC de Cavendisn". Sin datos propios y con "............ escasos conocimientos de química " (ditto Chargaff), este par de hiperactivos acomodan los componentes del DNA guiados por una lista de contundentes requisitos físicoquímicos establecidos previamente por otros investigadores (cuadro I). Los primeros modelos fallaron estrepitosamente. Uno, por crasa ignorancia de la química (usaron tautómeros equivocados). Otro, por intentar mono y trihélices antes de arribar en la DOBLE hélice, ésta basada evidentemente en la espléndida fotografía 51 de Rosalind Franklin, del DNA más hidratado, el tipo B, que llegó a sus manos en forma todos lo sabemos un tanto heterodoxa, por decirlo discretamente (Figura 8).

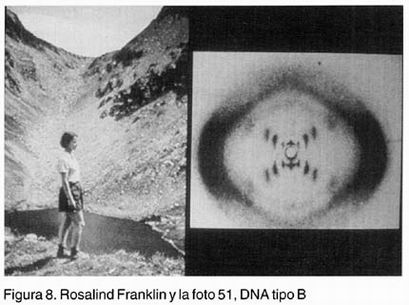
Para fortuna de Watson y Crick, el noruego Sven Furberg -que ya tenía un DNA monohélice plausible-abandona la carrera y se regresa a Oslo.Pauling por su parte, comete un error al construir laboriosamente una triple hélice , que no funcionó. Además el Departamento de Estado, en pleno macartismo, demora su viaje a Londres -por creerlo comunista- allá donde Franklin seguramente le hubiera mostrado de buena gana aquella foto de la forma B del DNA al gran señor de las hélices -y ya premio Nobel en 1954- antes que a ese par de locuaces advenedizos, que no hacían otra cosa sino discutir modelos interminablemente ........i........con datos ajenos! iPero qué soberbia discusión debe haber sido aquella. Y dos cerebros trabajan mejor que uno!
De no haber sido por estos incidentes, la loca carrera -"........ what mad persuit " (Crick) -de la doble hélice del DNA la hubiera ganado Pauling. ¿Lo habrán consolado con el Nobel de la Paz de 1962, año en que ya se dijo-Watson, Crick y Wilkins reciben el premio Nobel en Fisiología o Medicina?
Bibliografía
1. Benítez-Bribiesca L. DNA. The secret of life? Arch Med Res 2003;34:355. [ Links ]
2. Chargaff E. Heraclitan fire. New York: Rockefeller Univ Press; 1978. [ Links ]
3. Crick F. What mad pursuit. New York; Basic Books; 1988. [ Links ]
4. Dayson FJ. The sun, the genome and the Internet. Oxford, Univ. Press; 1999. [ Links ]
5. García Barreno P. 50 Años de ADN. La doble hélice. Madrid, España: Forum; 2003. [ Links ]
6. Judson HF. The eighth day of Creation. Cold Spring Harbor NY, USA: Cold Spring Harbor Press; 1996. [ Links ]
7. Magner LN. A history of life sciences. New York: M. Dekker; 1994. [ Links ]
8. Morange M. A history of molecular biology. Cambridge MA, USA: Harvard Univ. Press: 1998. [ Links ]
9. Murphy, NW, O'Neill, LAJ. What is Life? The next fifty years. Cambridge, UK: Cambridge Univ Press; 1995. [ Links ]
10. Portugal FH, Cohen JS. A century of DNA. Cambridge, MA. USA; MIT Press; 1979. [ Links ]
11. Schrödinger E. What is Life? New York: Cambridge Univ. Press; 1944. [ Links ]
12. Watson JD. The double helix. New York: Simon and Schuster; 1968. [ Links ]
13. Watson JD. Passion for DNA. Cold Spring Harbor, NY, USA: CSBL Press; 2000. [ Links ]

