











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Algunos autores documentan que las poblaciones y especies de plantas han desarrollado mecanismos de respuesta a señales ambientales que estimulan el desarrollo de las estructuras vegetativas y reproductivas (Hamann, 2004; Elzinga et al., 2007), y que en su conjunto generan el comportamiento a nivel de comunidad. Los detonantes ambientales a los que responden los patrones de floración y fructificación de los taxones arbóreos son variados y diferentes en función del ecosistema de que se trate; pues mientras que en las zonas templadas, las especies responden a factores evidentes como el fotoperiodo y la temperatura (Tooke y Battey, 2010; Diez et al., 2012), en las regiones tropicales lo hacen a la variación de la humedad atmosférica y del suelo, así como a la incidencia de la radiación solar (Hamann, 2004; Ochoa et al., 2008).
La presencia del agua, a través de la precipitación pluvial, ejerce una fuerte influencia sobre la fenología en los ecosistemas tropicales, pero su efecto tiene distinto grado de importancia en aquéllos con alto contenido de humedad y en los secos; además, es relevante cómo se distribuye la humedad en el año, lo cual genera comportamientos fenológicos diferenciados en los ecosistemas (Acevedo y Hernández, 2013).
Hay coincidencia en señalar que procesos evolutivos complejos han llevado a que las especies coincidan en sus tiempos de floración, en respuesta a factores ambientales que incrementen las probabilidades de su reproducción; entre ellos, factores abióticos como la precipitación, temperatura, fotoperiodo y factores bióticos como la presencia de polinizadores, herbívoros, etc. (Janzen, 1967).
En los bosques neotropicales estacionales secos y húmedos, se observa una tendencia a la concentración de la riqueza y abundancia de la floración de las especies arbóreas en la época de estiaje o periodos de menor humedad (Janzen, 1967; Williams y Meave, 2002; Cortés-Flores et al., 2019), patrón que también se registra en México para algunos bosques mesófilos de montaña (Hernández y Carreón, 1987; Solórzano et al., 2010); aunque para algunos bosques templados neotropicales el fotoperiodo es uno de los elementos de mayor influencia en la fenología floral de los taxones leñosos (Cortés-Flores et al., 2015).
La producción de frutos, se espera que ocurra cuando se presentan las mejores condiciones de humedad en el suelo para las diásporas, lo cual propicia que las semillas germinen y crezcan lo suficiente en su parte subterránea y aérea que les permita su supervivencia durante la primera temporada seca del año (van Schaik et al., 1993; Hamann, 2004; Zimmerman et al., 2007). Algunos patrones observados en la fructificación a nivel de comunidad, en los bosques tropicales secos, es que un conjunto de especies fructifican en la interfase de la estación seca-lluvias (Smythe, 1970), las cuales germinan en la estación lluviosa inmediata (taxones con semillas recalcitrantes); mientras que en otras, la fructificación ocurre al final de la temporada de lluvias y sus diásporas, las cuales, por lo general, tienen mecanismos de latencia de meses (especies con semillas ortodoxas), germinan hasta la siguiente temporada húmeda o en las posteriores (González et al., 2016; Topete et al., 2020).
En bosques tropicales, con poca o nula estacionalidad, los patrones de floración y fructificación de las especies arbóreas parece responder a la incidencia de radiación solar (Hamann, 2004; Zimmerman et al., 2007); mientras que, en los bosques templados mexicanos se observa que la longitud del día tiene un efecto positivo sobre la fenología floral (Cortés-Flores et al., 2015).
Entender el comportamiento de la floración y fructificación en los diferentes ecosistemas contribuye a explicar los patrones que rigen la distribución y abundancia de grupos de animales (Newstom et al., 1994); además, es de relevancia para determinar la época óptima de recolección de diásporas con fines de propagación (Brennan, 1996; Ochoa et al., 2008).
El propósito de la presente investigación fue describir la fenología reproductiva, floración y fructificación, a nivel de comunidad, de las especies arbóreas existentes en una zona montañosa del Occidente de México con alta heterogeneidad ambiental; para ello, se atendieron las siguientes preguntas: ¿Cómo varía la floración y fructificación de las especies arbóreas dentro del ciclo anual a nivel de comunidad?; ¿Existen diferencias en los patrones de floración y fructificación entre las comunidades vegetales existentes y con la altitud del terreno?; ¿Existe alguna correlación entre los patrones de floración y fructificación con la temperatura, precipitación, evaporación, humedad relativa y la radiación solar?
Materiales y Métodos
Área de estudio
La investigación se realizó en una zona montañosa de la Sierra Madre del Sur, un área que comprende un cuadrángulo de 6 × 6 km, con una superficie de 3 600 ha y un gradiente altitudinal de 1 500 a 2 250 msnm. El cuadrángulo se localiza entre los 19°34'14"-19°37'30" N y 104°14'49"-104°18'16" O, en los municipios Autlán de Navarro y Cuautitlán de García Barragán del estado de Jalisco; y dentro de sus límites se ubica la Estación Científica Las Joyas (ECLJ) (Jardel et al., 2004). La ECLJ es un predio propiedad del Gobierno del estado de Jalisco, forma parte de la zona núcleo Manantlán-Las Joyas en la Reserva de la Biosfera Sierra de Manantlán, Jalisco (Jardel et al., 2004). El centro de la ECLJ se sitúa 22 km al sur-sureste de la ciudad de Autlán de la Grana, 17 km al este de Casimiro Castillo y 52 km al norte del puerto de Manzanillo.
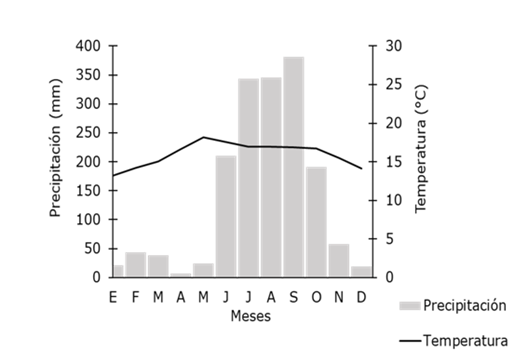
La vegetación corresponde a bosque mesófilo de montaña (BMM), bosque de Pinus (BP), bosque de Quercus (BQ), bosque de galería (BG), bosque de Pinus-Quercus (BPQ), vegetación secundaria (VS), renuevo de pino (RP), pastizal y agricultura (PA) y afloramiento de rocas y erosión (RE) (Jardel et al., 2004). Desde 1985, la ECLJ ha estado exenta de disturbios como la agricultura, ganadería y aprovechamientos forestales; sin embargo, eventualmente ocurren algunos incendios superficiales y la incidencia de ciclones como Jova y Patricia en el 2011 y 2015, respectivamente. El clima es marcadamente estacional, con una estación fría que comprende los meses de octubre a enero, una seca de febrero a mayo y una lluviosa, de junio a septiembre. Con los registros climáticos de los últimos 30 años, la media anual de temperatura y precipitación han sido de 15.5 °C y 1 663 mm, respectivamente; de acuerdo con información de la estación metereológica del lugar (Figura 1).
Obtención de los datos
Los especímenes disponibles en herbarios han sido fuente de información para estudios fenológicos (Borcher, 1996; Solórzano et al., 2010; Tooke y Battey, 2010). En el caso de la ECLJ y áreas aledañas, se ha realizado una recolección continua de material botánico desde 1984 hasta la fecha, material que forma parte del acervo del Herbario ZEA del Centro Universitario de la Costa Sur, y que representa una colección de especies arbóreas cercana a los 3 000 ejemplares. En la ECLJ también se tiene una estación climatológica, ubicada a 1 850 m de altitud, en la cual desde 1993 se registra información continua del área. Tanto las recolectas de ejemplares, como la información climatológica en el área permite, con cierto grado de confianza, realizar una investigación como la que se presenta en este documento.
Se llevó a cabo una revisión exhaustiva de los especímenes de herbario de especies arbóreas recolectadas en la ECLJ y áreas aledañas. Se consideraron como individuos en floración, los ejemplares con flores bien desarrolladas, con pétalos abiertos y anteras expuestas (Tooke y Battey, 2010); mientras que, los individuos en fructificación fueron aquellos con frutos bien desarrollados, maduros o muy cercano a ello. Con la información disponible en las etiquetas de los ejemplares de herbario se integró una base de datos mensual en Excel, que incluyó: género, especie, familia, fecha de recolecta, presencia de flor o fruto, tipo de vegetación (BMM, BP, BPQ, BQ, BG, VS) y altitud. Se consideraron los tipos de vegetación indicados, porque son los referidos en las etiquetas de los ejemplares de herbario. Los ejemplares cuyas recolectas presentaban flores, frutos o ambos y que se realizaron durante la primera semana o última del mes, se incluyeron en el mes previo o posterior, para tener una mejor representación en los periodos fenológicos. La nomenclatura de las especies se basó en la publicación de Cuevas et al. (2021).
Análisis de datos
La base de datos en Excel se pasó al programa InfoStat, en el cual se hicieron los análisis estadísticos (Di Rienzo et al., 2019). La estadística descriptiva se efectuó mediante histogramas. Se aplicaron análisis de normalidad de los datos con la prueba de Shapiro-Wilks y las pruebas de homogeneidad de varianza a través del procedimiento de Kolmogorov Smirnoff (Di Rienzo et al., 2019). Para analizar la variación de la floración y fructificación de las especies arbóreas se utilizaron análisis de varianza de una vía. Se realizó una prueba de significancia de Duncan a posteriori entre pares de medias para determinar las diferencias entre ellas, se consideró como significativa una p≤0.05. Para el análisis de los patrones de floración y fructificación, se consideraron tres estaciones: la fría que comprendió de octubre a enero, la seca de febrero a mayo y la estación de lluvias, de junio a septiembre.
Se calculó un promedio mensual de los valores de las variables climáticas registradas en la estación meteorológica de la ECLJ como insolación promedio (hr dia-1), temperatura media (°C), humedad relativa (%), evaporación (mm) y precipitación (mm). La relación entre cada una de estas variables y el número de especies en floración y fructificación se obtuvo por medio de correlaciones de Pearson.
Resultados
Floración y fructificación de las especies arbóreas a nivel de comunidad
De los 3 000 ejemplares examinados, solo 1 724 reunieron los criterios para considerarse en floración o fructificación, de acuerdo con lo definido en el método. Se registraron 124 especies, 96 géneros y 49 familias de especies arbóreas. Las familias con más ejemplares fueron Fagaceae con 181 (10.50 %), Solanaceae 130 (7.54 %), Fabaceae 102 (5.92 %), Malvaceae 88 (5.10 %) y Betulaceae 74 (4.29 %). La floración de los taxones arbóreos del área de estudio presentó un patrón de distribución bimodal, con un pico de floración de marzo-mayo y otro de noviembre-enero; con un notable decremento en el número de especies de agosto a octubre, aunque hubo taxones floreciendo todo el año (Figura 2). En relación con la fructificación, presentó su mayor riqueza de febrero a julio, con una disminución de agosto a noviembre, pero durante el año siempre hubo taxones en fructificación (Figura 2).

La riqueza de especies en floración fue mayor en la estación seca, no hubo diferencias estadísticas significativas con la estación fría, aunque sí con la estación lluviosa (F= 13.02, gl=2, p≤0.0022) (Figura 3a). La mayor riqueza de especie en fructificación se presentó en la estación seca y fue estadísticamente diferente a la fría y la lluviosa (F= 5.46, gl= 2, p≤0.0280) (Figura 3b).

Patrones de floración y fructificación por comunidades vegetales
El BMM fue la comunidad con mayor registro de ejemplares en floración y fructificación; y donde se identificó la riqueza más grande de familias, géneros y especies; mientras que, la menor riqueza de taxones y de ejemplares se observó en el BG (Cuadro 1).
Cuadro 1 Registro de ejemplares por comunidades vegetales y su riqueza de taxones.
| Vegetación | Eflores | Efrutos | Especies | Géneros | Familias |
|---|---|---|---|---|---|
| BG | 94 | 77 | 41 | 38 | 28 |
| VS | 101 | 92 | 46 | 39 | 26 |
| BP | 127 | 148 | 63 | 48 | 31 |
| BPQ | 159 | 118 | 64 | 49 | 26 |
| BQ | 161 | 168 | 76 | 54 | 32 |
| BMM | 397 | 383 | 115 | 91 | 47 |
BG = Bosque de galería; VS = Vegetación secundaria; BP = Bosque de Pinus; BPQ = Bosque de Pinus-Quercus; BQ = Bosque de Quercus; BMM = Bosque mesófilo de montaña (BMM); Eflores = Ejemplares registrados con flores; Efrutos = Ejemplares registrados con frutos.
El patrón de floración por comunidades vegetales fue el registrado para las especies de árboles en su conjunto. Hay una concentración de la riqueza de especies floreciendo para los meses de febrero a mayo y otra de noviembre a diciembre, con algunas pequeñas variantes en el BP, en el cual la floración presentó un notorio incremento de diciembre a marzo; mientras que, en el BQ el primer pico de floración se observó de marzo a abril y el segundo de noviembre a enero (Figura 4).

VS = Vegetación secundaria; BQ = Bosque de Quercus; BPQ = Bosque de Pinus-Quercus; BP = Bosque de Pinus; BMM = Bosque Mesófilo de Montaña; BG = Bosque de galería.
Figura 4 Patrones de floración por comunidades vegetales.
La fructificación por comunidad vegetal es un poco más heterogénea en su comportamiento, tiende al patrón que presentó la fructificación de todas los taxones. Se observó una mayor riqueza de especies en fructificación de febrero a julio, con una diminución de agosto a octubre y un incremento en noviembre y diciembre (Figura 5).

Patrones de floración y fructificación en relación a variables ambientales
Los patrones de floración estuvieron correlacionados en forma negativa y estadísticamente significativa con la humedad relativa (r = -0.91 p≤0.0001) y la precipitación (r = -0.79 p≤0.0003). No se obtuvo una relación significativa con la temperatura, evaporación y la insolación (Cuadro 2). La fructificación, sin ser significativa evidenció una fuerte correlación negativa con la humedad relativa (r =-0.58 p≤0.0561) y también una disminución en la riqueza de especies en fructificación con el incremento de la precipitación (Cuadro 2).
Cuadro 2 Correlaciones entre los patrones de riqueza de especies en floración y fructificación; riqueza de especies por comunidades vegetales con variables ambientales.
| Temperatura | Evaporación | Precipitación | Humedad relativa |
Insolación | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| r | p | r | p | r | p | r | p | r | p | |
| Flor | -0.40 | 0.194 | 0.16 | 0.628 | -0.79 | 0.0003 | -0.91 | ‹0.0001 | -0.06 | 0.8523 |
| Fruc | 0.15 | 0.636 | 0.37 | 0.242 | -0.41 | 0.187 | -0.58 | 0.0561 | 0.33 | 0.2889 |
| Floración por comunidad vegetal | ||||||||||
| VS | -0.34 | 0.277 | -0.01 | 0.984 | -0.72 | 0.0078 | -0.75 | 0.0046 | -0.16 | 0.6118 |
| BQ | -0.52 | 0.084 | -0.03 | 0.933 | -0.81 | 0.0015 | -0.88 | 0.0002 | -0.31 | 0.3293 |
| BPQ | -0.52 | 0.086 | -0.02 | 0.945 | -0.82 | 0.0010 | -0.90 | 0.0001 | -0.26 | 0.4189 |
| BP | -0.60 | 0.038 | -0.01 | 0.987 | -0.91 | ‹0.0001 | -0.97 | ‹0.0001 | -0.25 | 0.4357 |
| BMM | -0.36 | 0.247 | 0.14 | 0.655 | -0.74 | 0.0063 | -0.84 | 0.0006 | -0.04 | 0.9079 |
| BG | -0.40 | 0.204 | 0.30 | 0.346 | -0.55 | 0.0664 | -0.86 | 0.0004 | 0.10 | 0.7674 |
| Fructificación por comunidad vegetal | ||||||||||
| VS | -0.17 | 0.598 | 0.05 | 0.868 | -0.05 | 0.8688 | -0.12 | 0.7100 | 0.26 | 0.4233 |
| BQ | 0.01 | 0.970 | 0.23 | 0.474 | -0.19 | 0.5602 | -0.29 | 0.3566 | 0.36 | 0.2544 |
| BPQ | -0.35 | 0.262 | -0.53 | 0.075 | -0.29 | 0.3545 | 0.06 | 0.8531 | -0.33 | 0.2973 |
| BP | -0.17 | 0.605 | -0.08 | 0.808 | -0.19 | 0.5630 | -0.07 | 0.8238 | 0.12 | 0.7052 |
| BMM | -0.12 | 0.713 | 0.36 | 0.256 | -0.47 | 0.1261 | -0.58 | 0.0469 | 0.31 | 0.3284 |
| BG | -0.37 | 0.231 | 0.20 | 0.527 | -0.69 | 0.0127 | -0.82 | 0.0012 | 0.08 | 0.8028 |
Flor = Riqueza de especies en floración; Fruc = Riqueza de especies en fructificación; VS = Vegetación secundaria; BQ = Bosque de Quercus; BPQ = Bosque de Pinus-Quercus; BP = Bosque de Pinus; BMM = Bosque mesófilo de montaña; BG = Bosque de galería; r = Correlación; p = Valor de p.
Los patrones de floración presentaron una correlación negativa y significativa con la humedad relativa y la precipitación en las comunidades vegetales, con excepción del BG en donde se registró una disminución de la riqueza de especies en floración; aunque la correlación no fue significativa (Cuadro 2). La fructificación solo mostró una correlación negativa y significativa en el BG con la precipitación; y en el BMM y BG con la humedad relativa (Cuadro 2).
Patrones de floración y fructificación a través del gradiente altitudinal
La riqueza de especies en floración y fructificación fue mayor entre los 1 700 y 2 000 m de altitud, con una caída muy notoria arriba de los 2 100 m (Figura 6).

Discusión
Algunos autores han documentado (Williams y Meave, 2002; Solórzano et al., 2010) las limitaciones del uso de ejemplares de herbario en los estudios de fenología reproductiva, entre ellas la falta de un registro de la intensidad de las fenofases, de las abundancias de los individuos en floración o fructificación, o bien esta es muy limitada y diferenciada entre las especies; además de que, se requiere de recolectas abundantes y continuas en un área a través de varios años para tener una buena representación de la fenología de las especies. Con excepción de las dos primeras limitaciones, la recolecta continua de material botánico por más de 30 años en la ECLJ y áreas aledañas permite tener un buen grado de confianza en los patrones de fenología reproductiva determinados a nivel de comunidad y por tipos de vegetación.
Las 124 especies de árboles registradas para la ECLJ representan 27 % de esta forma biológica en la sierra de Manantlán (Vázquez et al., 1995). Las familias con más ejemplares en floración o en fructificación correspondieron con aquellas de mayor riqueza de especies de árboles en la ECLJ (Cuevas et al., 2004; Cuevas et al., 2021).
El patrón de floración registrado en la ECLJ, con una mayor riqueza de especies arbóreas floreciendo en la temporada seca (Figuras 2, 3a), se ajusta a lo citado para bosques tropicales estacionales, húmedos y en bosques templados (Janzen, 1967; Reich y Borchert, 1984; Wright y van Schaik, 1994; Moore, 2008; Ochoa et al., 2008; Cortés-Flores et al., 2015), así como en bosques mesófilos de México (Hernández y Carreón, 1987; Solórzano et al., 2010). Patrón atribuible a que durante este período las especies están en menor actividad fotosintética, después de la temporada de lluvias previa que permitió la acumulación de reservas (Singh y Kushwaha, 2006); además, también es cuando se presenta menor incidencia de plagas por insectos (Wright y van Schaik, 1994).
Hay otro conjunto de factores que pudiesen influir, algunos de ellos señalados por otros autores (Janzen, 1967; Moore, 2008), entre ellos: la abundancia de algunos grupos de polinizadores, principalmente abejas; la carencia o disminución de hojas hace las flores más visibles a los polinizadores; las flores no caen, ni disminuyen su concentración de néctar por las lluvias. La importancia del grupo de los insectos como polinizadores parece tener sustento, ya que con base en la información disponible (Cuevas et al., 2021), en función de los tipos de flores, se sugiere que 74 % de las especies arbóreas del área presentan síndromes de polinización entomófila, 21 % anemófila y 5 % ornitófila.
La fructificación presenta su mayor riqueza de febrero a julio, coincidiendo con la mayor parte de la temporada de estiaje y el inicio de la temporada de lluvias, con una notoria disminución en los meses con mayor precipitación (Figuras 2, 3b); patrón previamente registrado (Hamann, 2004; Cornejo e Ibarra, 2007; Solórzano et al., 2010). Este se ha explicado como lógico, en el sentido de que la producción de frutos en la temporada seca e inicio de lluvias minimiza la exposición de las semillas a los depredadores y provee tiempo para que las diásporas germinen y el establecimiento de las plántulas ocurra con más éxito, ya que se aprovecha la humedad disponible en el ambiente durante el corto temporal de lluvias, sobre todo la del suelo (Hamann, 2004; Singh y Kushwaha, 2006).
Las especies que presentan sus picos de fructificación a finales del periodo de lluvias posiblemente tienen diásporas con periodos de latencia, lo cual permitiría que sean viables hasta la próxima temporada de lluvias.
Dado que la riqueza de especies en floración y fructificación en los meses con menos riqueza no es menor a 20 (Figura 2), a pesar de que el área presenta una clara temporada estacional de lluvias y secas, así como un periodo frío, se considera que hay condiciones de mayor humedad en el ambiente que las existentes en las regiones tropicales secas, lo cual genera condiciones para la presencia de especies con estructuras reproductivas a lo largo del año; esto coincide con lo registrado en otros trabajos para México, en los que la vegetación predominante es el bosque mesófilo de montaña (Hernández y Carreón, 1987; Solórzano et al., 2010).
Es importante considerar que en las comunidades vegetales del área, la presencia de materia orgánica en el suelo debe contribuir al mantenimiento de la humedad edáfica, lo cual genera condiciones para que las especies arbóreas sean perennifolias o caducifolias por periodos cortos, con la existencia de cierto grado de actividad fotosintética; por lo que se observan especies con flores y frutos durante todo el año.
El repunte en la floración y fructificación en la riqueza de especies que se observa de octubre a diciembre (Figura 2) podría corresponder con una disminución en la precipitación que inicia a partir de octubre (Figura 1). La disminución en la riqueza de especies en floración y fructificación en enero sería el resultado de una baja en la temperatura, o una menor incidencia de recolecta de ejemplares en ese mes.
La riqueza de especies en floración y fructificación fue mayor en el BMM (Figuras 4, 5; Cuadro 1), lo cual se esperaba, si se considera que en esta comunidad vegetal se registra la mayor riqueza de especies arbóreas (Cuevas et al., 2004; Cuevas et al., 2021). El patrón de la floración y fructificación es acorde con lo esperado y refleja el comportamiento de la fenología reproductiva de la flora arbórea completa de la ECLJ. Las explicaciones a este patrón serían las indicadas previamente para el conjunto de las especies de árboles del área.
La humedad relativa y la precipitación fueron los dos factores ambientales con mayor influencia sobre la floración, los cuales parecen ser señales a las que responden las especies disminuyendo su floración (Cuadro 2). La explicación para ello, es que estos factores ambientales tienen influencia tanto en las flores, como en la actividad de los polinizadores (Janzen, 1967). Mayor humedad y agua implican más dificultad para la actividad de los polinizadores, ya que se obstaculiza su movilidad; además hay una disminución en la recompensa de las flores al diluirse el néctar y disminuir su calidad nutricional; también, se reduce la cantidad de flores por impacto de las gotas y el incremento de su peso con el agua; asimismo hay más nubosidad y por consiguiente menos horas luz, lo cual disminuye sus actividades (Janzen, 1967).
Las variables temperatura, evaporación y la insolación parecen no fluctuar mucho en el área, y no mostraron relación significativa con la riqueza de floración, contrario a las evidencias registradas en otras regiones (Siegmund et al., 2016) (Cuadro 2).
La riqueza de especies en fructificación mostró una clara tendencia a disminuir con el incremento en la humedad relativa (p≤0.0561) y con el incremento de la precipitación, lo cual es coincidente con lo indicado en la literatura (Cornejo e Ibarra, 2007; Cortés-Flores et al., 2012); y que se atribuye al hecho de que las especies maximizan el tiempo de disponibilidad de humedad para que las diásporas germinen y tenga lugar el establecimiento de las plántulas (Janzen, 1967).
La riqueza de especies en floración y fructificación fue mayor entre los 1 700 y 2 000 m de altitud, con una disminución muy notoria arriba de los 2 100 m, lo cual responde a un efecto de área, ya que 67 % de la superficie de la ECLJ se ubica en el intervalo altitudinal de 1 700-2 000 m, y <1 % por arriba de los 2 200 m (Figura 6).
Conclusiones
Los patrones de floración y fructificación de las especies arbóreas del área de estudio son coincidentes con lo registrado para los bosques tropicales secos, los húmedos con cierto grado de estacionalidad en su precipitación y los bosques templados húmedos de México, así como en otras regiones de Centroamérica.
La condición de estacionalidad más importante a la que parecen responder la fenología reproductiva de los árboles de la ECLJ está determinada por la humedad relativa y la precipitación; pero a pesar de ello, siempre hay especies con flores o frutos todo el año.
En el área de estudio, factores ambientales como la temperatura, la insolación y la evaporación no varían mucho a través del año y sus efectos sobre los patrones de fenología reproductiva de las especies arbóreas parecen no tener mucha influencia.
Para un mayor entendimiento de los patrones fenológicos de las especies de plantas en la ECLJ es necesario analizar la fenología reproductiva de arbustos y herbáceas e incluir en el análisis los síndromes de polinización y dispersión de las especies.

