











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Uno de los pasos esenciales durante los proyectos de restauración es la selección de las especies a utilizar (Vargas Ríos, 2011). En México algunos aspectos tomados en cuenta durante la restauración de ecosistemas terrestres han sido, entre otros, favorecer la regeneración natural, así como la selección de material biológico nativo (Méndez-Toribio et al., 2018). Sin embargo, a pesar de esto las bajas tasas de supervivencia del material vegetativo integrado, después del primer año de haber sido plantado, empañan este tipo de opciones como las únicas preferentes.
Aunque existen diversos factores sociales y económicos que pueden mermar la persistencia del material vegetativo utilizado en programas de reforestación u otras acciones de restauración forestal, en el ámbito técnico las posibles razones de la baja supervivencia observada es la utilización de especies y procedencias inadecuadas, por lo que se ha sugerido diversificar las especies y procedencias producidas en viveros, así como establecer ciertos criterios encaminados a mejorar los niveles de ajuste de los genotipos a los ambientes, y realizar acciones encaminadas a mejorar la conservación de los recursos genéticos forestales (Sáenz-Romero & Linding-Cisneros, 2004).
En particular, preocupa la conservación de los recursos genéticos derivados del Pinus hartwegii. Esto está relacionado con diversos efectos causados por el cambio climático que han afectado, significativamente, a la distribución de hábitats adecuados para ésta, y otras especies de coníferas, situación que se según Gómez‐Pineda et al., (2020) afectará de forma más alarmante, hacia finales de este siglo.
En la actualidad se ha reportado que existe una importante diferenciación genética entre poblaciones de P. hartwegii, a lo largo de gradientes ambientales heterogéneos. En ese sentido, Viveros-Viveros et al., (2009) reportaron variación genética en el crecimiento de plántulas de esta especie, provenientes de poblaciones distribuidas a lo largo de un gradiente altitudinal en el Pico de Tancítaro, Michoacán. También, Loya-Rebollar et al., (2013) encontraron diferencias en varios caracteres entre procedencias, indicando la presencia de un patrón de diferenciación genética atribuido a la altitud, y por tanto al clima. Al respecto Aparicio-Rentería et al., (2020), plantean que resulta necesario realizar investigaciones sobre la variación de los atributos morfológicos (conos, semillas y acículas), ya que esto podría ayudar a delimitar zonas provisionales para el aprovisionamiento y movimiento de germoplasma.
El presente trabajo se centra en el estudio de la relación de caracteres fisiológicos de semillas y fenotipos de P. hartwegii asociados con un gradiente altitudinal útil para delimitar zonas preliminares de suministro de germoplasma, con el propósito de que esto sirva de referencia en las actividades de reforestación y conservación de recursos genéticos forestales. La investigación base que da origen al trabajo se realizó en la Estación Forestal Experimental Zoquiapan (EFEZ) de la Universidad Autónoma Chapingo, ubicada en el Estado de México. Tiene como objetivo determinar el nivel de variación de los caracteres fenotípicos y fisiológicos entre las procedencias (diferentes altitudes) de Pinus hartwegii para el tamaño de la semilla, tamaño del cono, longitud de la acícula, capacidad y velocidad de germinación de la semilla, y definir si existe un patrón de variación clinal para estos caracteres en la EFEZ. A la par, se buscó establecer zonas altitudinales para el suministro y movimiento de germoplasma según la variación clinal.
Métodos
Área de estudio
El área de estudio se estableció en la EFEZ, estación de investigación perteneciente a la Universidad Autónoma Chapingo (Fig. 1), ubicada en el municipio de Ixtapaluca, Estado de México, entre las coordenadas 19º 12' 30" y 19º 20' 00" de latitud norte y 98º 42' 30" y 98º 30' 00" de longitud oeste. Su clima es templado subhúmedo con lluvias en verano y estación seca en invierno; presenta una temperatura media anual de 11 ºC y una precipitación media anual de 964 mm. La vegetación predominante es bosque de pino, bosque de oyamel, bosque de aile y pastizales (Universidad Autónoma Chapingo, 2020).

Diseño de muestreo
Entre los meses de marzo y mayo de 2022, de forma aleatoria se recolectó material biológico consistente en cinco conos femeninos, maduros y sanos por árbol, de la misma manera se cortaron de la mitad inferior de copa cinco ramas con fascículos maduros (con un ciclo de crecimiento), de las cuales se eligió un fascículo por cada una, al que se le midió la acícula más larga, en un transecto altitudinal (de 3265 a 3650 msnm) que incluyó la variación existente entre las procedencias de Pinus hartwegii, en el polígono seleccionado. Se consideró como exposición cardinal promedio la sudeste y una zona en la cual la especie se distribuye de manera natural en el cerro nombrado localmente como “El Papayo”.
Con ayuda de un Sistema de Posicionamiento Global (GPS) (GARMIN, Map 78s), se ubicaron las procedencias recolectadas en promedio cada 50 m de altitud (Tabla 1). Se muestrearon ocho procedencias, excepto la procedencia de 3550 m de altitud, por tener baja densidad de árboles, pocos estróbilos femeninos maduros y ser de difícil acceso. En cada procedencia, se seleccionaron al azar de cinco a 10 árboles, con una distancia mínima de 30 m entre ellos para reducir la probabilidad de parentesco (Erickson & Adams, 1989).
Tabla 1 Localización geográfica en UTM de ocho procedencias muestreadas de P. hartwegii en la EFEZ, Ixtapaluca, Estado de México, México.
| Procedencia | Altitud (m) |
Latitud (norte) |
Longitud (oeste) |
No. de árboles |
|---|---|---|---|---|
| 1 | 3650 | 19.307481 | -98.698560 | 10 |
| 2 | 3600 | 19.306261 | -98.698033 | 7 |
| 3 | 3500 | 19.304712 | -98.697707 | 5 |
| 4 | 3450 | 19.303265 | -98.697050 | 9 |
| 5 | 3400 | 19.299849 | -98.694478 | 7 |
| 6 | 3350 | 19.297305 | -98.693561 | 6 |
| 7 | 3300 | 19.294050 | -98.683872 | 10 |
| 8 | 3265 | 19.290031 | -98.676982 | 7 |
Toma de datos
Variables morfológicas
Las variables morfológicas evaluadas fueron: longitud de conos, longitud de acículas, largo y ancho de semillas. Con la ayuda de un vernier digital (STEREN, HER-411) se midieron los conos desde la base hasta la punta; de esta forma se obtuvo la longitud de cada cono por árbol y por procedencia. Igualmente, con la ayuda de una regla graduada, se midió la acícula más larga de cada uno de los cinco fascículos de cada árbol y se ordenaron por procedencia. En cuanto a las semillas, en cada procedencia se recogió la semilla obtenida de sus respectivos árboles y se mezclaron las semillas de los árboles de cada procedencia. De ésta se seleccionaron 90 semillas llenas (obtenida a partir de flotación en agua, donde las semillas sumergidas se consideraron llenas y las que flotaron vanas) al azar de cada procedencia. Las semillas seleccionadas se midieron tanto en largo (de la base a la punta) como en anchura (parte más ancha) con la ayuda del vernier digital.
Prueba de germinación
La prueba de germinación se realizó en una cámara de germinación, acondicionada con bombillas eléctricas domésticas de diferentes voltajes para ajustar la temperatura a una media de 25 ºC ± 3 ºC. En recipientes de plástico de 30 cm de largo por 20 cm de ancho por 15 cm de alto se vertió perlita como sustrato, que fue previamente desinfectada con el fungicida-bactericida OxiDate 2.0® (BioSafe Systems, LLC, USA) a una dosis de 1 ml/l de agua, se agitó la solución con el sustrato y se dejó reposar durante 3 días con el recipiente tapado. La semilla se remojó durante 24 h en agua corriente y luego se desinfectó con el fungicida CAPTANE 50 PH® (Agroquímica Tridente S.A de C.V., México) a una dosis de 3 g/l de agua filtrada durante 30 minutos.
Se utilizó un total de 100 semillas por procedencia, mediante un Diseño experimental en bloques completos al azar, se consideró a 25 semillas en cada bloque una réplica, por lo que en total fueron cuatro bloques en los cuales la semilla se sembró al azar dentro de una franja (el bloque contuvo ocho franjas, una para cada procedencia).
El ensayo tuvo una duración de 40 días en los que la lectura de la germinación se realizó diariamente durante dos semanas (tiempo en el que se presentó una germinación constante), y posteriormente cada tercer día, ya que la germinación fue esporádica. El criterio para considerar una semilla germinada fue el agronómico (emergencia y desarrollo de una plántula) (Pita-Villamil & Pérez-García, 1998). Los datos de capacidad germinativa (%) fueron obtenidos con la siguiente fórmula, y la velocidad de germinación contando los días necesarios para alcanzar el 50% de la capacidad germinativa, ambos para cada procedencia.
Donde:
cg = capacidad germinativa en porcentaje.
sg = número de semillas germinadas.
ts = total de semillas sembradas.
Procesamiento y análisis de datos
Para análisis de los datos de las variables morfológicas, capacidad germinativa y velocidad de germinación se empleó el software estadístico SAS (versión 9.4; SAS Institute Inc., Cary, NC, USA). Para la velocidad de germinación se realizó una comparación de medias utilizando el programa IBM SPSS Statistics© versión 25.
Variables morfológicas
Para determinar la existencia de variación entre procedencias se realizó un análisis de varianza con el procedimiento Proc Mixed de SAS, para cada una de las variables fenotípicas (longitud de la acícula, longitud del cono, largo y ancho de la semilla), usando un nivel de significancia α = 0.05, donde el modelo utilizado fue:
Donde:
Yijk = valor de la ijk-ésima observación
µ = media general del experimento
Pi = efecto de la i-ésima procedencia
A(P)ij= efecto del j-ésimo árbol anidado en la i-ésima procedencia
εijk = término de error aleatorio
El mismo procedimiento se utilizó para obtener los componentes de la varianza en los efectos considerados como aleatorios (árbol anidado en la procedencia y error aleatorio). Posteriormente, para visualizar qué procedencias diferían entre sí, se realizó la comparación de medias con el método de Tukey-Kramer, usando en este caso un nivel de significancia de 0.2 (valor de α considerado para determinar la zonificación descrita posteriormente).
Diseño experimental para la prueba de germinación
El modelo estadístico de análisis utilizado fue:
Donde:
Yijk = valor de la ijk-ésima observación
µ = media general del experimento
Bi = efecto del i-ésimo bloque
Pj = efecto de la j-ésima procedencia (altitud)
εij = término de error aleatorio
Para estimar la variación entre procedencias se utilizó el procedimiento de SAS Proc Glimmix para la variable capacidad germinativa, y Proc Freq (para obtener la prueba no paramétrica de Friedman) para la variable velocidad de germinación usando un nivel de significancia de α = 0.05. Asimismo, para esta variable se realizó la prueba no paramétrica de Wilcoxon, que permitió la comparación de medias entre procedencias con un α = 0.1.
Posteriormente, para observar el posible comportamiento clinal a lo largo del transecto altitudinal, se realizaron diagramas de dispersión de los porcentajes medios de capacidad germinativa y de los valores medios de velocidad de germinación frente a sus respectivas altitudes de origen, ajustando un modelo de regresión a los datos según su significación (α = 0.05) y su coeficiente de determinación (r2).
Zonificación
Zonificación altitudinal
Para los análisis de varianza de las variables morfológicas en los que existió diferencia significativa entre procedencias se calculó la Diferencia Mínima Significativa, DMS (Least Significant Difference: LSD) entre medias correspondiente a un α = 0.2. Para observar si existía un patrón altitudinal de la variación entre procedencias, se realizó una regresión entre las medias por procedencia y su altitud de origen. Finalmente, con base en la variable largo de semilla se realizó una zonificación altitudinal provisional considerando un clima contemporáneo (promedio 1961-1990), basado en transferir la DMS entre procedencias a su equivalente en altitud y climática (Sáenz-Romero, 2004).
Zonificación para clima contemporáneo y clima futuro (RCP 6.0)
El clima contemporáneo (temperatura) de referencia utilizado fue el promedio de los años 1961 a 1990. El clima futuro fue el centrado en la década de 2060, estimado por un ensamble de modelos-escenarios, spline para todo Norte América, con una Vía de Concentración Representativa o RCP, con un forzamiento radiactivo de 6.0 W/m2. Ambos fueron obtenidos de la plataforma online de Investigación sobre el Cambio Climático Forestal: http://charcoal.cnre.vt.edu/climate/. La información proporcionada por esta plataforma se generó a partir de datos originados por el Laboratorio de Ciencias Forestales de Moscow del Servicio Forestal en Idaho, USA, del Departamento de Agricultura de los Estados Unidos, USDA.
Una vez obtenidos los datos climáticos puntuales, se repitió el procedimiento descrito en el apartado "Zonificación altitudinal", pero ahora se cambiaron los datos del eje de abscisas por la temperatura media anual del clima contemporáneo (1961-1990) correspondiente a cada una de las altitudes de origen, obteniendo así una zonificación para el clima contemporáneo. Para la zonificación del clima futuro, también se repitió el procedimiento, pero, los datos del eje de abscisas se cambiaron por la temperatura climática media anual estimada para la década de 2060 (Ortiz-Bibian et al., 2017; Sáenz-Romero et al., 2012).
Una vez definida la zonificación, se sugirieron directrices para la toma de decisiones en el movimiento de germoplasma dentro de las zonas y entre ellas, según se consideraran o no los efectos del cambio climático (Loya-Rebollar et al., 2013).
Resultados
Variables morfológicas
El análisis de la varianza indicó que de las cuatro variables analizadas sólo el largo de semilla mostró diferencia significativa entre procedencias a un α = 0.05. (Tabla 2) muestra los P-valores correspondientes al ANOVA de cada variable.
Tabla 2 P-valores asociados al análisis de varianza para procedencias originadas a diferentes altitudes, considerando una significancia de 0.05 y porcentaje de contribución a la varianza total.
| Variables | P-valor | Porcentaje de contribución a la varianza total (%) | ||
|---|---|---|---|---|
| Procedencia | Árbol (Procedencia) | Error | ||
| Longitud de cono | 0.1054 | 6.49 | 56.61 | 36.89 |
| Longitud de acícula | 0.0910 | 8.25 | 62.52 | 29.23 |
| Ancho de semilla | 0.2671 | 2.56 | 57.06 | 62.52 |
| Largo de semilla | 0.0058 | 16.82 | 44.46 | 38.71 |
La comparación de pares de medias entre procedencias para el largo de semilla indicó que las procedencias ubicadas aproximadamente en la mitad del transecto altitudinal difieren en el valor promedio con respecto a las procedencias ubicadas en los extremos del mismo transecto (Tabla 3), no existiendo diferencias entre las demás comparaciones de pares de medias.
Tabla 3 Comparación de Procedencias originadas a diferentes altitudes que resultaron significativamente diferentes para el Largo de semilla según la prueba de Tukey-Kramer (α = 0.2).
| Altitud de origen (msnm) / media ajustada (mm) | P-valor | |
|---|---|---|
| 3650 / 4.49 | 3400 / 5.09 | 0.1793 |
| 3650 / 4.49 | 3350 / 5.33 | 0.0205 |
| 3400 / 5.09 | 3265 / 4.37 | 0.0834 |
| 3350 / 5.33 | 3265 / 4.37 | 0.0089 |
Germinación
El factor procedencia tuvo un efecto altamente significativo (p < 0.0001) para la variable respuesta capacidad germinativa. En el análisis de regresión, se observó un patrón de variación cuadrático (significativo con la omisión de una procedencia atípica) de los valores medios de germinación contra su altitud de origen (r2 = 0.7865; p = 0.0456), reforzando así la existencia de variación clinal en la especie para el largo de la semilla.
Sin embargo, para la velocidad de germinación se encontró una débil variación significativa (p = 0.0556) entre procedencias. La comparación de pares de medias indicó que existe una débil diferencia estadística entre las procedencias 4 (3450 m) y 3 (3500 m) con una p = 0.059, 8 (3265 m) y 3 (3500 m) con p = 0.066, 5 (3400 m) y 4 (3450 m) con p = 0.066 y, 8 (3265 m) y 7 (3300 m) con p = 0.068, pero sin un patrón aparente de distribución altitudinal. En la Tabla 4 se muestran los valores obtenidos para las variables fisiológicas de la semilla.
Zonificación altitudinal
El valor de la DMS obtenida considerando un alfa de 0.2 para la variable largo de semilla fue de 0.27 mm, que por razones prácticas se redondeó a 0.3 mm. El modelo de regresión cuadrática (r2 = 0.88, p = 0.0144) entre la media por procedencia para largo de semilla contra su altitud de origen fue el que mejor se ajustó, sin tener en cuenta una altitud atípica (por tener un desempeño muy distinto a la tendencia general) a 3350 msnm (Fig. 2).
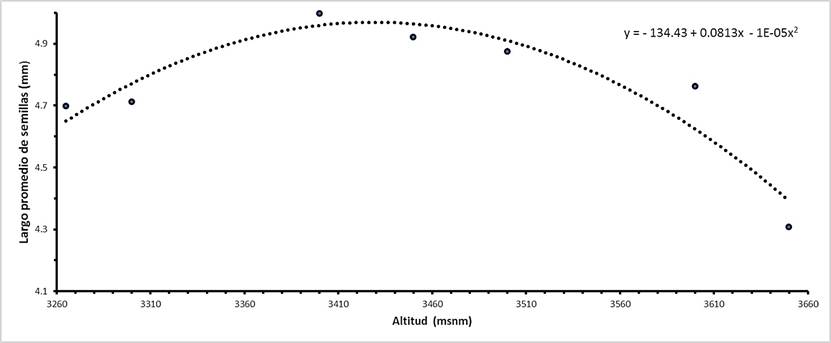
Fig. 2 Comportamiento del largo promedio de semilla de P. hartwegii respecto a la altitud de origen de siete procedencias en la EFEZ.
Posteriormente, a partir del valor de la DMS, se delimitaron tres zonas altitudinales para el aprovisionamiento y movimiento de germoplasma a lo largo del transecto altitudinal. Debido a que la zona altitudinal más alta cubría un rango muy pequeño (aproximadamente 70 m de diferencia altitudinal), se decidió fusionar las tres zonas en sólo dos zonas, partiendo de una altitud inferior fija de 3250 m como punto de partida. Este punto inicial arbitrario sigue perteneciendo a la distribución altitudinal natural de la especie (Tabla 5).
Tabla 5 Límites altitudinales y climáticos de dos zonas fijas para el aprovisionamiento y movimiento de germoplasma con clima contemporáneo.
| Zona | Elevación (m) | Zona | Media anual de temperatura (ºC) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Límite (m) | Rango (m) | Intervalo (±) | Límite (ºC) | Rango (ºC) | Intervalo (±) | ||||
| Inferior | Superior | Inferior | Superior | ||||||
| 1 | 3250 | 3450 | 200 | 100 | 1 | 8.0 | 9.0 | 1.0 | 0.5 |
| 2 | 3450 | 3650 | 200 | 100 | 2 | 7.0 | 8.0 | 1.0 | 0.5 |
Zonificación para clima contemporáneo y clima futuro (RCP 6.0)
Bajo el mismo razonamiento de simplificación realizado para la zonificación en base a la altitud, para la zonificación climática, la zona número tres (parte más alta del transecto altitudinal) tenía un rango muy estrecho (0.3 ºC), por lo que, se decidió fusionarlas dejando solo dos zonas (ver Tabla 5).
En cuanto a la zonificación para el clima proyectado al año 2060, comparando con los límites de las zonas fijas con el clima contemporáneo, se estimó a qué altitudes se tendrían que desplazar las poblaciones para reacoplarse a las temperaturas para las cuales están adaptadas actualmente, pero que ocurrirán a mayor altitud debido al cambio climático. Se estimó que las temperaturas que conforman los límites de las zonas fijas de germoplasma se desplazan altitudinalmente hacia arriba aproximadamente 400 m (Fig. 3).

En consecuencia, se esperaría que en aproximadamente cuatro décadas (década centrada en 2060), las zonas climáticas fijas actuales estarían fuera de los límites altitudinales del cerro "El Papayo" (Tabla 6).
Tabla 6 Límites altitudinales y climáticos para dos zonas fijas de germoplasma considerando un clima proyectado al año 2060, RCP 6.0.
| Zona | Elevación (m) | Media anual de temperatura (ºC) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Límite (m) | Rango (m) | Intervalo (±) | Límite (ºC) | Rango (ºC) | Intervalo (±) | ||||
| Inferior | Superior | Inferior | Superior | ||||||
| 12060 | 3650 | 3850 | 200 | 100 | 8.0 | 9.0 | 1.0 | 0.5 | |
| 22060 | 3850 | 4050 | 200 | 100 | 7.0 | 8.0 | 1.0 | 0.5 | |
Lineamientos para el movimiento de germoplasma
Las directrices para el abastecimiento y movimiento de germoplasma que se utilizará en actividades de reforestación o restauración ecológica, con base en lo planteado por Loya-Rebollar et al., (2013) serían:
1) Sin considerar los inminentes efectos del cambio climático:
a) Obtención de germoplasma de una de las zonas fijas, producir plantas en vivero y plantarlas dentro de la misma zona (delimitada ya sea por intervalos altitudinales o climáticos) donde se recolectó el material de propagación.
b) Recolectar germoplasma a una altitud cualquiera dentro del rango natural de distribución de la especie en la región, producir plantas y plantarlas a intervalos de ± la mitad del ancho de una zona fija (ver Tabla 5); es decir, ± 100 m hacia arriba o hacia abajo con respecto a la altitud de recolección o ± 0.5 º C con respecto a la temperatura media anual del punto de recolección (zonas de punto focal).
2) Si se tienen en cuenta los efectos del cambio climático en la dinámica de las poblaciones de P. hartwegii, se debe considerar que las temperaturas actuales a las que se establecen naturalmente las poblaciones se moverán aproximadamente 400 m más arriba (ver Figura 3). Así, los límites climáticos que determinan las zonas fijas para determinar el sitio de plantación se situarán fuera del límite altitudinal del cerro "El Papayo" (3650 m).
3) Si se considera que los intervalos climáticos y altitudinales de las zonas futuras2060 son los mismos que los de las zonas contemporáneas, y con el objetivo de realinear poblaciones de P. hartwegii a un clima al que actualmente están adaptadas, pero que debido al calentamiento global se espera su desplazamiento a 400 m de altitud más arriba, se recomienda:
a)Recolectar germoplasma en una zona contemporánea, producción de planta en vivero y plantarla a 400 m de altitud en promedio más arriba, lo que implica recolectar germoplasma de la zona 1 contemporánea y plantar en la zona 1 futura del año 2060; para la zona 2 contemporánea se debe repetir el proceso descrito para la zona 1.
Cabe mencionar que si las zonas climáticas fijas contemporáneas se desplazaran altitudinalmente fuera de los límites altitudinales del cerro "El Papayo", aún existe la posibilidad de que en el futuro existan condiciones de temperatura adecuadas en el área para las poblaciones de P. hartwegii que actualmente se desarrollan a altitudes menores a las consideradas en el transecto altitudinal utilizado en este estudio, y para las cuales podrían transponerse los límites altitudinales o de zona climática aquí delimitados.
Discusión
Variables morfológicas
El largo promedio de semilla (4.9 mm) encontrado para P. hartwegii en este estudio es similar al reportado por Iglesias et al., (2012) y Niembro-Rocas (1992) de 4.9 mm y valores entre 3.4 a 5.4 mm respectivamente. En cuanto al ancho promedio de semilla (3.5 mm), éste fue similar al reportado por Iglesias et al., (2012) de 3.4 mm. Asimismo, estuvo dentro del rango de valores reportado por Niembro-Rocas (1992) de 2 mm a 3.7 mm.
La longitud promedio del cono (8.59 cm) fue similar al mostrado por Iglesias et al., (2012) de 7.82 ± 1.1 cm. Asimismo, estuvo dentro del rango de valores descrito por Iglesias-Andreu & Tivo-Fernández (2006) quienes reportaron valores entre 4.2 a 11.7 cm para esta característica. Por su parte, Farjon et al., (1997) los situaron en el rango de 8 a 12 cm.
En cuanto a la longitud promedio de la acícula más larga (18.63 cm) fue ligeramente superior al descrito en Guízar-Nolazco (2014) quien indica valores entre 10 y 17 cm, y contrastado drásticamente por lo publicado por Iglesias-Andreu & Tivo-Fernández (2006) quienes indican valores medios para la especie, pero en poblaciones del Cofre de Perote en Veracruz, México de 8.4 ± 1.71 cm, aunque Farjon et al. (1997) indican valores de hasta 22 cm de longitud.
En cuanto a la variación morfológica de caracteres con posible valor adaptativo como los estudiados, se plantea que se debe a la plasticidad fenotípica o a la adaptación a diferentes condiciones ambientales o incluso a ambas, como se observó en un estudio realizado en México, relacionado con la variación del tamaño de las hojas para Quercus rugosa Nee (Uribe-Salas et al., 2008). Al respecto, se considera que esto podría explicar la variación encontrada en el tamaño de las acículas en el presente estudio con respecto a otros publicados sobre esta especie.
Ahora bien, la diferenciación encontrada entre procedencias para el largo de semilla en P. hartwegii, también fue reportada por Iglesias et al., (2012) entre las poblaciones del Cofre de Perote y Pico de Orizaba en el estado de Veracruz, México. Cabe decir, a diferencia de ellos en el presente estudio, no se encontraron diferencias significativas en las variables longitud de acícula más larga y longitud de cono (p ≤ 0.05).
Germinación
La existencia de diferencias significativas en la capacidad germinativa entre las procedencias de P. hartwegii encontradas en este estudio también se han reportado en otras especies de coníferas. Al respecto Ortiz-Bibian et al. (2019) para Abies religiosa, en Michoacán, México, donde a lo largo de un gradiente altitudinal las procedencias del centro presentaron mayor capacidad germinativa y viabilidad; coincidiendo con este estudio para la capacidad germinativa (donde las procedencias de 3400 y 3500 msnm presentaron un 90% y 78% de germinación media respectivamente y una velocidad de germinación media de 13 y 13.25 días, respectivamente), siendo las procedencias situadas en los extremos del transecto las de menor porcentaje de germinación media. Estos autores también coinciden con el patrón de variación altitudinal encontrado para la capacidad de germinación, sugiriendo que se debe a las condiciones ambientales que prevalecen en los extremos altitudinales de distribución de la especie, como estrés por heladas (en la parte alta) y eventualmente sequía (en la parte baja).
Montero-Nava (2020) también reportó los valores más altos de germinación en P. hartwegii a una altitud de 3400 m (89%), para un transecto altitudinal en Puebla, indicando que los valores de capacidad germinativa disminuían a medida que aumentaba la altitud a partir de esta ubicación (55% a 4000 m), lo que también coincide con lo hallado en el presente estudio (tendencia de menor capacidad germinativa promedio).
Cabe mencionar que en el presente estudio se observó que las procedencias de los extremos altitudinales presentaban una menor densidad de árboles de la especie en estudio, lo que podría repercutir en el proceso de polinización, y, por lo tanto, en el desarrollo de las semillas, así como en el proceso de germinación, aunque esto debe tomarse con reserva ya que no se realizaron correlaciones con la densidad de árboles.
Zonificación altitudinal y climática
El patrón de variación altitudinal del largo promedio de semilla, indica la existencia de una tendencia (cuadrática) específicamente para el cerro "El Papayo". Según López-Ríos (2009) el óptimo ecológico existente para las especies forestales se sitúa en la parte media de la distribución altitudinal, por lo que hablando de temperatura los valores ecológicos máximos y mínimos se situarían en los extremos altitudinales inferior y superior, respectivamente.
En cuanto a la temperatura se considera que tiene una alta correlación negativa con la altitud, (Fig. 3).
Teniendo en cuenta que los factores ambientales pueden influir en el desarrollo de los individuos en función de la capacidad genética que poseen los taxones en los rangos de tolerancia, se podría inferir que la plasticidad fenotípica de P. hartwegii se expresa en este caso, influenciada por la variación de la temperatura a lo largo del gradiente altitudinal, un patrón que ha sido reportado para árboles (Thomas, S. C. 2011), y que posiblemente denote que existe una diferenciación genética entre las procedencias atribuida a la presión de selección ejercida por el mismo cline de temperatura a lo largo del gradiente altitudinal, aunque, no es el único aspecto que influye, ya que hay más factores además de la temperatura que podrían estar interviniendo.
Sin embargo, esto último tendría que ser verificado con pruebas que incluyan rasgos cuantitativos con valor adaptativo como lo hicieron Loya-Rebollar et al. (2013), quienes determinaron diferencias genéticas entre poblaciones de P. hartwegii para diferentes rasgos como: alargamiento total, tasa de alargamiento, altura final de la plántula, diámetro basal, peso seco de acículas, ramas, tallo y peso seco total; mismos que se agruparon en un componente principal y reflejaron un patrón de variación (modelo cuadrático) a lo largo de un gradiente climático-altitudinal.
Asimismo, se reportó que la temperatura media anual, entre otros elementos a considerar, fue una variable climática que resultó significativa debido a que permite predecir los valores medios por población del primer componente principal antes mencionado, lo que coincide con lo encontrado en este trabajo, con un modelo cuadrático (r2 = 0.84, p = 0.0251) para el largo promedio de semilla por procedencia frente a la temperatura media anual.
Thomas S.C. (2011) indica que las especies arbóreas comúnmente presentan cambios morfológicos y fisiológicos dramáticos cuando se distribuyen a lo largo de gradientes altitudinales, siendo la disminución de la temperatura y su relación con el aumento de altitud el factor que posiblemente más impacta directamente a los procesos fisiológicos de las plantas, ya que con el aumento de altitud se ha encontrado un importante incremento en valores de la fotosíntesis saturada de luz, la conductancia estomática, el nitrógeno foliar, así como la masa foliar por área, y comúnmente el tamaño de las hojas disminuye.
Estos clines climáticos-altitudinales significativos han sido reportados para diferentes especies de coníferas en atributos morfológicos y rasgos cuantitativos, tales como Pinus devoniana en Michoacán (Castellanos-Acuña et al., 2013; Sáenz-Romero et al., 2012), Abies religiosa (Castellanos-Acuña et al., 2014; Ortiz-Bibian et al., 2017, 2019), P. patula en Oaxaca (Viveros-Viveros et al., 2013) y en Veracruz (Aparicio-Rentería et al., 2020).
En este sentido, el rango de altitud y temperatura media anual correspondiente a cada una de las dos zonas delimitadas en este estudio coincidió con el determinado por Loya-Rebollar et al. (2013) para la misma especie, y fue el mismo que se especificó por Ruiz-Talonia et al. (2014) en P. patula, para la altitud. En ambos casos, las zonificaciones se basaron en la diferenciación genética de rasgos cuantitativos, lo cual es similar a lo planteado por Castellanos-Acuña et al., (2014) para A. religiosa (250 m).
Lineamientos para movimiento de germoplasma
Una premisa aceptada por la comunidad de investigadores forestales es que la adaptación de las poblaciones arbóreas a ambientes heterogéneos (tanto en el espacio como en el tiempo) depende de la variación genética que éstas posean, así como de la capacidad que tienen para modificar el fenotipo (a nivel individual) en relación con el ambiente (Chambel et al., 2005). En ese sentido, se considera que cuando el material de propagación disminuye su capacidad de adaptación al enfrentar determinadas condiciones desfavorables presentes en los lugares de plantación, se da una alta mortalidad antes de que los individuos alcancen la edad reproductiva; así como un crecimiento reducido y un descenso en la producción de semillas, e incluso la pérdida de competitividad entre individuos (Thomas et al., 2014).
Bajo el supuesto de que las poblaciones naturales de árboles ocupan su espacio de distribución actual porque están adaptadas a las condiciones ambientales del mismo, como resultado de un largo proceso evolutivo, entonces las directrices sobre el movimiento de germoplasma, sin considerar el cambio climático dentro de la misma zona fija o considerando las zonas de puntos focales, son opciones que se ajustarían al supuesto, ya que se consideran las zonas que proporcionan a las poblaciones arbóreas las condiciones a las que están adaptadas. En consecuencia, el establecimiento de individuos bien adaptados en actividades de reforestación o restauración es más probable que ocurra con más éxito en cada zona, por lo que su uso puede ser una herramienta útil en la gestión forestal.
Por otro lado, no se pueden ignorar los efectos inminentes y potencialmente desastrosos del cambio climático sobre las poblaciones de árboles. En ese sentido, se ha previsto un aumento del fenómeno de la sequía, que afecta el funcionamiento fisiológico de los árboles y por consiguiente los debilita (Del-Val & Sáenz-Romero, 2017; Gómez-Pineda et al., 2022). En el mismo sentido, el aumento proyectado de temperatura media anual promedio para la zona de estudio fue de 2.3 ºC (año 2060), lo que puede en un futuro, dificultar los procesos de establecimiento y crecimiento de las plantas, ya que de acuerdo con E. Thomas et al. (2014), la presión de selección aumenta cuando el clima es más inhóspito.
La estimación mundial del aumento de la temperatura y su relación con la desviación del lugar de distribución de las poblaciones de varias especies arbóreas ha generado incertidumbre sobre su adaptación a temperaturas más cálidas (Hammond et al., 2022). En México, las coníferas no están exentas de esta situación, en ese sentido, Gómez‐Pineda et al. (2020) reportaron que para finales de siglo (año 2090) especies como P. oocarpa sufrirán una reducción de su hábitat climático propicio de 67%, P. devoniana de 58%, Abies religiosa de 88%, P. hartwegii de 87% con respecto a su área de distribución actual (año 1961-1990) a nivel nacional. Así, para P. hartwegii se prevé que para el año 2060 su clima favorable desaparecerá en varias de las montañas donde se distribuye actualmente, y éste aparecerá en montañas que superan los 4300 msnm, como el Nevado de Colima, Nevado de Toluca, Popocatépetl, Iztaccíhuatl, Pico de Orizaba y el Cofre de Perote.
Otros autores como Manzanilla-Quiñones et al. (2019) reportaron proyecciones de incrementos de temperatura media anual para el Eje Neovolcánico Transversal para el periodo 2041-2060 de entre 0.7 ºC y 1.5 ºC, con pérdidas del bosque subalpino de P. hartwegii de entre 11.7% y 18.3% y, 1.1 ºC y 2.1 ºC con pérdidas de entre el 18.3% y el 26.9% respecto a su distribución actual, lo que sugiere que para el mismo periodo de tiempo la especie se vería obligada a ascender altitudinalmente entre 100-500 m, lo que coincide con los hallazgos de este estudio, con un desplazamiento de la especie de aproximadamente 400 m para que las poblaciones actuales se vuelvan a acoplar a las condiciones de temperatura a las que están adaptadas.
La migración natural de las poblaciones de algunas especies arbóreas a mayores alturas como posible efecto del cambio climático se ha dado por sentada en México, por ejemplo, para el oyamel (Romahn-Hernández et al., 2020). En el caso de P. hartwegii, Alfaro-Ramírez et al. (2017) informan que no está claro si la especie podrá migrar altitudinalmente por encima de su límite altitudinal superior, debido a las condiciones ambientales adversas a las que está expuesta la especie en esas alturas.
Sin embargo, aunque las poblaciones pueden migrar altitudinalmente, las tasas de migración pueden ser superadas por las tasas de aumento de la temperatura (Peñuelas et al., 2007), por lo que la migración asistida puede servir como una herramienta con el propósito de reacoplar las poblaciones con sus climas favorables que en el futuro aparezcan en altitudes mayores. Este tipo de acciones han sido probadas en lugares como Canadá (Marris, 2009)) y han sido sugeridas en otros lugares para diferentes especies (Carbajal-Navarro et al., 2019; Loya-Rebollar et al., 2013); donde el uso de plantas nodriza como las del género Lupinus en sitios de reforestación a mayor altitud podría ser una estrategia para ayudar al establecimiento de individuos procedentes de áreas fijas ubicadas a menor altitud (Ramírez-Contreras & Rodríguez-Trejo, 2009).
Conclusiones
En la Estación Forestal Experimental Zoquiapan, México, se observa una diferenciación fenotípica (rasgos con posible valor adaptativo) para el largo de semilla, que mostró diferencias entre las procedencias por su altitud dentro de esta población de P. hatwegii originadas a lo largo de su transecto natural de distribución altitudinal, donde las procedencias centrales difieren significativamente de las ubicadas en los extremos del transecto climático-altitudinal.
La presencia de un patrón de distribución clinal significativo en los atributos fenotípicos (largo de semilla y capacidad germinativa) de P. hartwegii con respecto al transecto climático-altitudinal en el cerro “El Papayo”, puede ser utilizado para la delimitación de zonas fijas para el aprovisionamiento y movimiento de germoplasma en actividades de reforestación local.
Se sugiere el uso de las zonas delimitadas con un ancho de 200 m altitudinales o de 1 ºC, para actividades de recolección de germoplasma, sin considerar al cambio climático. También, se sugiere un movimiento altitudinal de 400 m hacia arriba de las fuentes de semilla, considerando el cambio climático proyectado al año 2060.
El aumento previsto de la temperatura media anual de 2.3 ºC para la década centrada en el año 2060 en la zona de estudio, indica que las poblaciones de P. hartwegii, siempre y cuando exista el espacio y las condiciones del suelo lo permitan, necesitarán ayuda para migrar altitudinalmente hacia zonas más altas. Esto no significa necesariamente la desaparición de la especie en la zona, pero sí una mayor presión de selección para las poblaciones actuales que podrían: a) desarrollarse en condiciones menos favorables, con el costo que ello conlleva: menor crecimiento y mayor susceptibilidad al ataque de plagas y enfermedades, o b) la presión de selección eventualmente dejaría sólo a los individuos con la constitución genética adecuada para superar las nuevas condiciones.

