











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Para formar híbridos comerciales competitivos de maíz (Zea mays L.) se requiere seleccionar poblaciones con media y varianza genética altas en las características agronómicas de interés (Rebolloza et al., 2016) para producir líneas endogámicas que al combinarse generen híbridos con la máxima expresión heterótica en la característica deseada.
Comstock et al. (1949) generaron el método de mejoramiento interpoblacional de selección recurrente recíproca (SRR) que consiste en seleccionar dos poblaciones divergentes genéticamente, A y B, donde las familias seleccionadas de la población A se cruzan con la población B como probador y viceversa, se evalúan las cruzas y las familias con mayor aptitud combinatoria general (ACG) se recombinan y se inicia un nuevo ciclo de selección; de esta forma, se capitalizan los efectos aditivos en las poblaciones y los no aditivos en la cruza.
Por su estructura, la SRR es el método genotécnico más utilizado para formar híbridos de maíz, al cual se le han hecho modificaciones para mejorar la eficiencia en la selección de líneas y cruzas (Paterniani y Vencovsky, 1978; Ramírez-Díaz et al., 2015; Souza Jr., 1987). Los resultados demuestran que la SRR ha sido efectiva para modificar la ACG de las poblaciones y la aptitud combinatoria específica (ACE) de sus cruzas en rendimiento de grano y otras características agronómicas (Keeratinijakal y Lamkey, 1993; Moll y Hanson, 1984; Romay et al., 2011; Santos et al., 2007). No obstante, existen resultados donde una de las poblaciones del par heterótico no presenta ganancia genética en rendimiento de grano u otras características agronómicas a través de los ciclos de selección (Moll y Hanson, 1984; Romay et al., 2011); es posible que la falta de ganancia genética se deba a que se recombinaron progenitores con valores aditivos bajos en características muy correlacionadas con rendimiento de grano o que no haya buena ACE para rendimiento de grano entre los progenitores seleccionados, pero esto no puede probarse porque se desconoce la ACE. Otra causa es la endogamia que se genera cuando se utilizan pocas líneas para formar las poblaciones (Astete y de Miranda, 2002; MárquezSánchez, 2013).
Una opción al problema anterior sería seleccionar de una a tres líneas más en la prueba de mestizos, hacer cruzas dialélicas (Griffing,1956) y confirmar el valor aditivo y no aditivo de las líneas seleccionadas, para recombinar sólo progenies con alta ACG y ACE y eliminar aquellas que contribuirán negativamente en la población. Otras ventajas de combinar ambas pruebas serían obtener cruzas simples con alta ACE para usarse como probadores de híbridos trilineales y dobles al combinarse con líneas y cruzas simples derivadas de la población recíproca usada como probador. Con el propósito de comprobar la estrategia propuesta, esta investigación tuvo como objetivo seleccionar, en una población de maíz subtropical, líneas y cruzas con alta ACG y ACE para rendimiento de grano y características agronómicas, al combinar la prueba de mestizos y de cruzas dialélicas
MATERIALES Y MÉTODOS
Material genético
En el Programa de Mejoramiento Genético de Maíz del Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias (INIFAP) se trabaja con el par heterótico Sintético Tardío (ST) y la Población de Amplia Base Genética Tardía (PABGT-CE). La población ST se formó con seis líneas élite S3 , cuatro tropicales y dos subtropicales, de la cual se seleccionaron los progenitores B-39 y B-41 que participan en los híbridos comerciales H-318, H-377
(Ramírez et al., 2010), H-516 (Gómez et al., 2001) y H-155, entre otros. La PABGT-CE se integró con 20 familias de medios hermanos maternos de germoplasma subtropical adaptado, seleccionadas por su resistencia al carbón de la espiga [Sphacelotheca reiliana (Khun) Clint] en Zapopan, Jalisco en condiciones de humedad residual (Ramírez y Ron, 1988).
Método de selección
Se utilizó el método de SRR, con la modificación de recombinar los progenitores seleccionados en mestizos después de haberse evaluado en cruzas dialélicas; esta modificación requiere de un ciclo agrícola más para completar un ciclo de SRR, independientemente del tipo de familia que se utilice como unidad de selección, pero tendría las ventajas siguientes: a) podría resolver el problema de la pérdida de ganancia genética en rendimiento de grano en algunas de las poblaciones del par heterótico (Moll y Hanson, 1984; Romay et al., 2011), porque se recombinarían familias con ACG y ACE altas, y b) en las cruzas dialélicas se identificarían líneas y cruzas simples que pueden usarse como probadores de cruzas trilineales y dobles, lo cual es importante para el INIFAP porque los usuarios directos de sus híbridos son empresas nacionales productoras de semilla que demandan semilla registrada de este tipo de híbridos para aumentar la rentabilidad en la producción de semillas.
En este método de mejoramiento se usaron líneas S3 como unidad de selección, pues la correlación genética entre una línea S3 y su cruza de prueba es 0.93 (Bernardo, 1991), valor alto que da confianza para seleccionar líneas y cruzas para formar híbridos comerciales de maíz. En la Figura 1 se presenta el esquema de mejoramiento; nótese que sólo se describen los pasos para una población del par heterótico, pero lo mejor es que el esquema se aplique simultáneamente en ambas poblaciones.
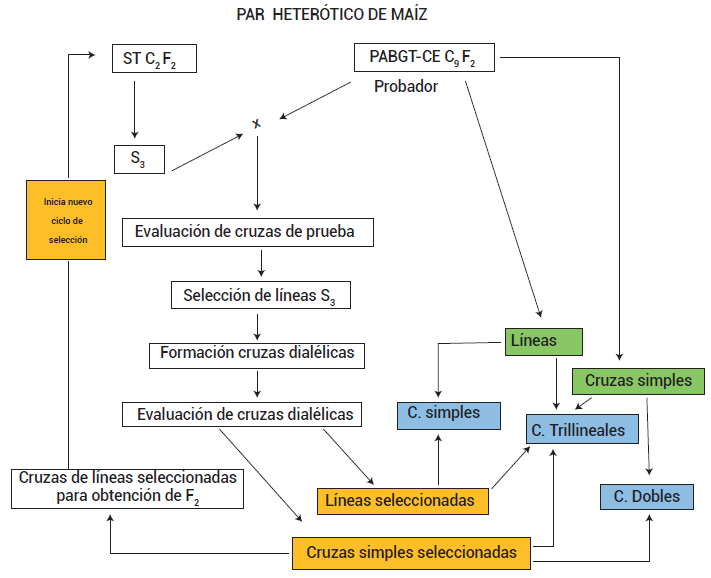
Figura 1. Esquema de selección del par heterótico ST C2 F2 × PABGT-CE C9 F2 que asocia la prueba de mestizos con cruzas dialélicas entre las líneas seleccionadas en la población. En el esquema se asume que las líneas y cruzas seleccionadas en la población PABGT-CE siguieron el mismo proceso de selección que en la población ST.
Formación de mestizos
En el ciclo agrícola de otoño-invierno (OI) 2010-2011 se sembraron, en condiciones de riego en terrenos del Campo Experimental Santiago Ixcuintla del INIFAP, 148 líneas S3 de la población ST C2 F2 y el probador población PABGT-CE C9 F2 para formar mestizos. Las líneas y el probador se sembraron en aislamiento en proporción 2:1 (líneas:probador). El tamaño de parcela fue un surco de 4.0 m distanciados a 0.80 m con 22 plantas cada uno. El manejo agronómico del lote se hizo con base en las recomendaciones para el cultivo del maíz generadas por el Programa de Maíz del Campo Experimental Santiago Ixcuintla (INIFAP, 2017a). En la etapa de floración, las espigas de las líneas se eliminaron antes de que iniciara la liberación de polen, y en la cosecha se eliminaron mazorcas de plantas acamadas y enfermas.
Evaluación de mestizos
En el ciclo agrícola de primavera-verano (PV) 2012 se integró un ensayo uniforme con 144 mestizos, las poblaciones originales, siete testigos comerciales del INIFAP y tres de empresas privadas. El ensayo se sembró en condiciones de temporal en las localidades de San Miguel Cuyutlán, municipio de Tlajomulco, Jalisco, ubicado a 1350 msnm, Peña del Panal, Michoacán (1897 msnm), Tepatitlán, Jalisco (1930 msnm) y San José de Mojarras, Nayarit (1084 msnm). Se utilizó el diseño experimental de bloques completos al azar con dos repeticiones. El tamaño de parcela fue de un surco de 4.0 m de longitud distanciados a 0.80 m, con 24 plantas cada uno, equivalente a la densidad de población de 75 mil plantas ha-1. El manejo agronómico de los experimentos se hizo con base en las recomendaciones técnicas para el cultivo del maíz generadas por el INIFAP en cada localidad (INIFAP, 2017a ; 2017b; 2017c).
Las variables estudiadas fueron a) rendimiento de grano en peso seco (kg ha-1) el cual se estimó con la ecuación siguiente:
Rendimiento = PC × PG/100 × PMS × FC
donde PC: peso de campo en mazorca (kg), PG: porcentaje de grano, PMS: (100 - porcentaje de humedad del PC)/100, FC: factor de conversión = 10,000/área de la unidad experimental = 3125.
Se evaluó además, b) número de días a floración masculina, contados desde la fecha de siembra hasta queen el 50 % de las plantas las espigas estuvieran liberando polen, c) número de días a floración femenina, contados desde la fecha de siembra hasta que el 50 % de las plantas exhibieron jilotes con estigmas expuestos con longitud mayor que 3.0 cm, d) altura de planta (cm), medida en tres plantas por parcela desde el suelo hasta la punta de la espiga, e) altura de mazorca (cm), medida en tres plantas por parcela desde el suelo hasta el nudo de la mazorca principal, f) porcentaje de acame de raíz, se calculó a partir del número de plantas acamadas de raíz dividida entre el número total de plantas de cada parcela (NTPL) y multiplicada por 100, se consideraron plantas acamadas de raíz aquellas desviadas 30o de la vertical o con “cuello de ganso”, y g) porcentaje de acame de tallo, calculado a partir del número de plantas acamadas de tallo dividida entre el NTPL y multiplicada por 100, se consideraron plantas acamadas de tallo a aquellas dobladas o quebradas abajo del nudo donde se inserta la mazorca principal.
Se hicieron dos tipos de análisis de varianza combinados, uno que incluyó sólo a los mestizos y el otro a los mestizos y testigos; se consideró a las localidades como efectos fijos y a los genotipos como aleatorios. En la comparación de medias se aplicó la prueba de la Diferencia Mínima Significativa (P ≤ 0.05) (SAS Institute, 1996). La aptitud combinatoria general (ACG) de los mestizos se calculó como la diferencia entre el valor de la variable obtenida en el mestizo y la media general de los mestizos (Hallauer y Miranda, 1988). La prueba estadística de ACG del mestizo se hizo dividiendo el valor de la ACG entre la desviación estándar de la media (S/√n) (Hallauer y Miranda, 1988), donde S es la raíz cuadrada del cuadrado medio del error y n es el número de localidades; éste valor se comparó con el valor crítico obtenido en la tabla de t, prueba de dos colas (P ≤ 0.05) y los grados de libertad del error, el cual fue igual a 1.97.
Formación y evaluación de cruzas dialélicas
Con base en el análisis conjunto de la prueba de mestizos, del 20 % de mestizos sobresalientes se seleccionaron nueve por rendimiento de grano y tolerancia al acame de raíz y tallo, principalmente. Con la semilla remanente de las nueve líneas seleccionadas en PV 2015, en la localidad de San Miguel Cuyutlán municipio de Tlajomulco, Jalisco, se hicieron las cruzas dialélicas con base en el modelo IV de Griffing (1956), del que se obtuvieron 36 cruzas simples.
En el ciclo agrícola PV 2016 las cruzas simples se evaluaron en un ensayo uniforme integrado con 36 cruzas simples, las poblaciones originales, cuatro testigos comerciales del INIFAP y dos de empresas privadas. Se utilizó el diseño experimental látice triple 7 × 6. El tamaño de parcela y la densidad de población fueron iguales a las usadas en la prueba de mestizos. El ensayo se sembró en cinco localidades, las cuatro ya señaladas para mestizos, más Iguala, Guerrero (751 msnm), de las cuales, cuatro se sembraron en temporal, y en Peña del Panal, Michoacán en punta de riego. El manejo agronómico de los experimentos se hizo con base en las recomendaciones técnicas para el cultivo del maíz generadas por el INIFAP en cada localidad (INIFAP, 2017a; 2017b; 2017c; 2017d). Las variables estudiadas fueron las mismas descritas en la evaluación de mestizos.
En el análisis estadístico combinado sólo se incluyó a las cruzas dialélicas. En la comparación de medias se aplicó la prueba de la DMS (P ≤ 0.05), y el análisis genético de las cruzas dialélicas se realizó con el programa propuesto por Zhang y Kang (2003). Los análisis estadísticos se hicieron utilizando el paquete estadístico de SAS® (SAS Institute, 1996).
RESULTADOS Y DISCUSIÓN
Evaluación de mestizos
En el análisis estadístico conjunto que incluyó mestizos, poblaciones originales y testigos, hubo diferencias significativas (P ≤ 0.05) para todas las variables, excepto altura de planta, en el factor variedades × ambiente. Las medias de rendimiento de grano del 20 % de mestizos sobresalientes fueron estadísticamente iguales a las de los testigos INIFAP-4 (7,545 kg ha-1) y TC-1 (6,736 kg ha-1), lo cual indica que su rendimiento de grano es competitivo (Cuadro 1).
Cuadro 1. Rendimiento de grano y características agronómicas del grupo sobresaliente (20 %) de mestizos formados con líneas S3 de la población ST C2 F2 usando como probador la población PABGT-CE C9 F2 . Promedio de cuatro localidades. 2012.
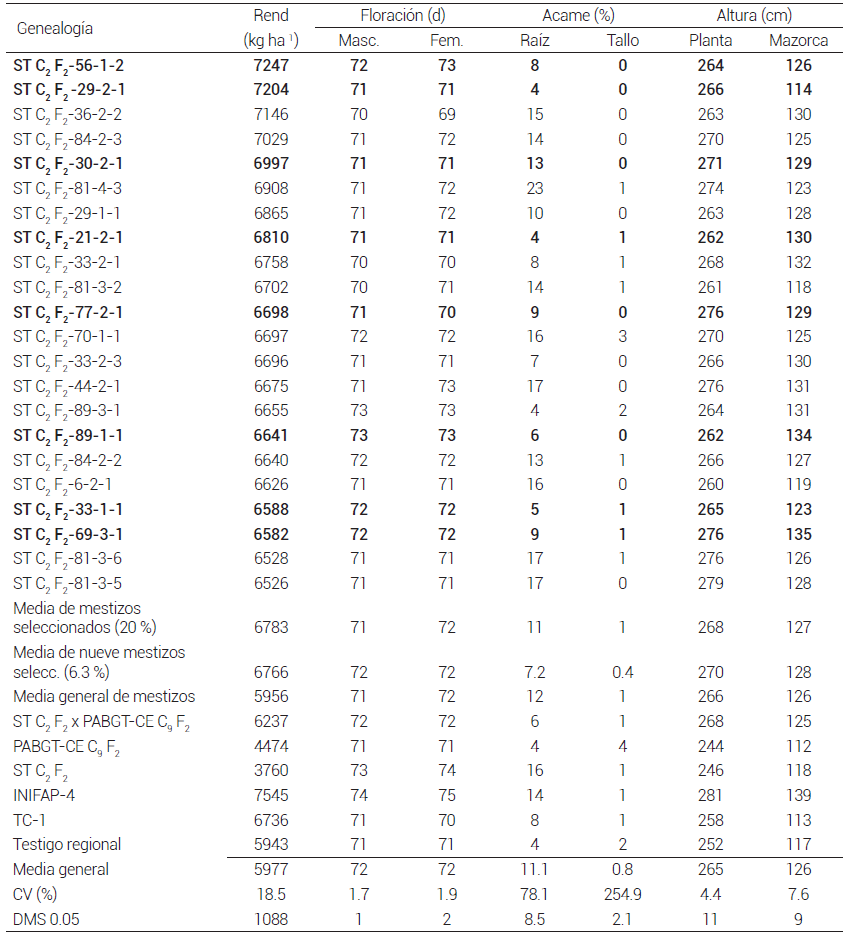
Rend:rendimiento de grano. En negritas los mestizos seleccionados para el siguiente ciclo de selección.
Por otra parte, en el análisis estadístico conjunto que incluyó sólo mestizos, hubo diferencias estadísticas significativas (P ≤ 0.05) para todas las variables en las fuentes de variación, excepto para altura de planta y mazorca en la interacción mestizos x ambientes. Los resultados anteriores indican diferencias entre ambientes y un comportamiento diferencial de los mestizos a través de los ambientes de evaluación (Cuadro 2)
Cuadro 2. Cuadrados medios del análisis de varianza de mestizos formados con líneas S3 de la población ST C2 F2 . Análisis combinado a través de cuatro localidades. 2012.
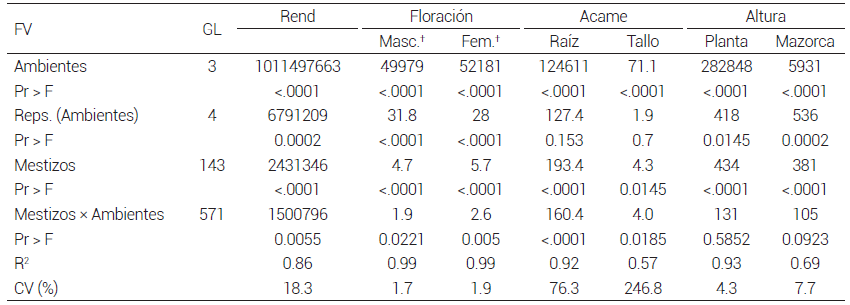
†Las variables floración masculina y femenina sólo se evaluaron en tres localidades por lo que los grados de libertad correctos para ambientes, reps. (ambientes), mestizos y mestizos × ambientes son 2, 3, 143 y 286, respectivamente. FV: fuente de variación, GL: grados de libertad, Rend: rendimiento de grano.
La media más alta de rendimiento de grano en localidades se registró en Tlajomulco (7761 kg ha-1) y fue mayor (P ≤ 0.05) que en el resto de localidades, le siguieron las localidades de Morelia (7525 kg ha-1), Santa María del Oro (5019 kg ha-1) y Tepatitlán (3812 kg ha-1), y entre éstas hubo diferencias significativas (P ≤ 0.05) (Cuadro 3). El rendimiento de grano más bajo obtenido en Tepatitlán se debió a una fuerte sequía que se presentó en la etapa final de llenado de grano, que pudo comprobarse con la media de altura de planta y mazorca, pues éstas fueron mayores (P ≤ 0.05) que en el resto de las localidades lo que indicó que el desarrollo de las plantas fue normal hasta la etapa reproductiva inicial. Al respecto, Çakir (2004) señala que la falta de humedad en el maíz en las etapas de antesis y llenado de grano reduce entre 66 y 93 % el rendimiento de grano. Por otro lado, la media más alta de acame se registró en Tlajomulco, debido a que el 28 de agosto de 2012 hubo una tormenta con vientos fuertes que acamó las plantas (Cuadro 3).
Cuadro 3. Rendimiento de grano y características agronómicas de mestizos formados con líneas S3 de la población ST C2 F2 y como probador la población PABGT-CE C9 F2 , en cuatro localidades. 2012.
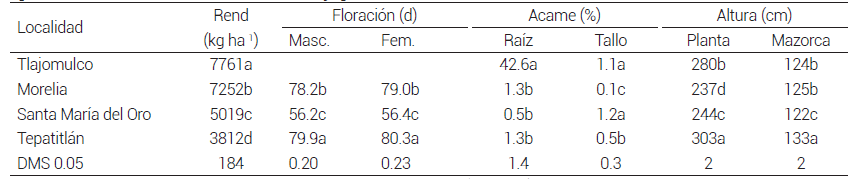
Medias con la misma letra en las columnas no son estadísticamente diferentes (DMS, 0.05). Rend: rendimiento de grano.
Inicialmente, se aplicó un 20 % de presión de selección en los mestizos evaluados, cuya media de rendimiento de grano, con respecto a la de la población, tuvo un diferencial de selección de 827 kg ha-1. Asimismo, la media del porcentaje de acame de tallo se redujo 8.3 % y la de acame de raíz fue igual a la de la población original. Considerando que la tolerancia al acame es una característica muy importante en los híbridos comerciales por la cosecha mecanizada, dentro del grupo del 20 % sobresaliente se seleccionaron nueve mestizos cuyo diferencial de selección en rendimiento de grano fue de 810 kg ha-1, 17 kg menos que cuando se seleccionó el 20 % de la población, pero el porcentaje de acame de raíz y tallo se redujeron 37 y 43 % con respecto al de la población original (Cuadro 1).
Se considera que nueve líneas es un número adecuado para integrar el sintético del siguiente ciclo de selección, pues Córdova y Márquez (1979) no encontraron diferencias en rendimiento de grano en sintéticos formados entre cinco y ocho líneas, y Márquez-Sánchez (2013) determinó que en un sintético de maíz formado con 10 líneas S1 y con la participación de 10 plantas por familia, la endogamia estimada fue baja (F= 0.0526).
De los nueve mestizos seleccionados, los formados con las líneas ST C2 F2 -56-1-2 y ST C2 F2 -29-2-1 tuvieron los valores aditivos positivos y significativos (P ≤ 0.05) más altos para rendimiento de grano. En floración masculina el valor aditivo del mestizo con la línea ST C2 F2 -89-1-1 fue positivo y significativo (P ≤ 0.05), lo que contribuiría a hacer más tardía la población, mientras que en floración femenina el valor aditivo del mestizo de la línea ST C2 F2 -77-2-1 fue negativo y significativo (P ≤ 0.05), el cual aportaría precocidad a la población. En altura de planta y mazorca, el mestizo de la línea ST C2 F2 -49-2-1 tuvo valores aditivos positivos y significativos (P ≤ 0.05) y es el que contribuiría más en modificar estas características en la población, mientras que el valor aditivo del mestizo de la línea ST C2 F2 -29-2-1 fue negativo y significativo (P ≤ 0.05), lo que contribuiría a reducir la altura de mazorca en la población. En las variables porcentaje de acame de raíz y tallo, en ninguno de los mestizos seleccionados hubo valores aditivos significativos, pero en la mayoría los valores fueron negativos, por lo que se esperaría que la población mejore en la tolerancia al acame (Cuadro 4).
Cuadro 4. Aptitud combinatoria general de mestizos formados con líneas S3 de la población ST C2 F2 y como probador la población PABGT-CE C9 F2 . Promedio de cuatro localidades. 2012.
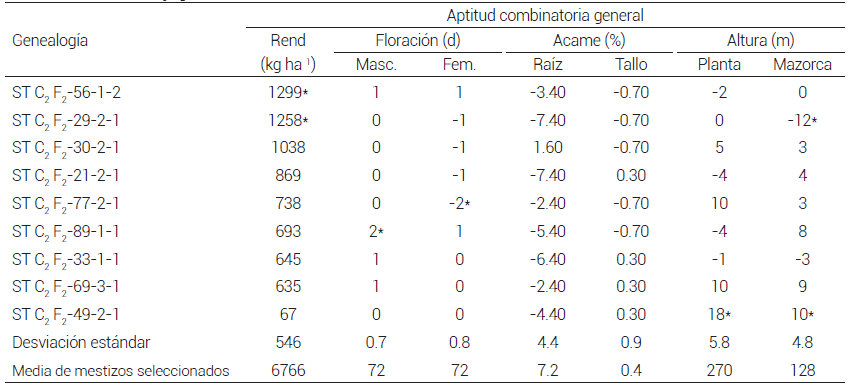
*Valor significativo (P ≤ 0.05)
El rendimiento de grano per se y la tolerancia al acame de la población PABGT-CE C9 F2 (4,474 kg ha-1 y 8 %) fueron mejores que en la población ST C2 F2 (3,760 kg ha-1 y 17 %). La heterosis interpoblacional, con respecto al progenitor medio y mejor progenitor, fue 51.5 % y 39.4 %, respectivamente, valores que están dentro de los encontrados por Keeratinijakal y Lamkey (1993), quienes en maíz encontraron que la heterosis del C0 al C11 varió de 25.4 a 76 % después de 11 ciclos de SRR.
En el método original de SRR (Comstock et al., 1949), una vez terminada la selección de cruzas se continuaría con la recombinación de progenies seleccionadas, obtención de las generaciones F1 y F2 del sintético seleccionado, y se iniciaría un nuevo ciclo de selección. En esta investigación el objetivo fue recombinar sólo progenies con valores aditivos y no aditivos altos en rendimiento de grano y características agronómicas para integrar el nuevo sintético, y además, seleccionar líneas y cruzas simples élite como probadores de híbridos simples, trilineales y dobles; entonces, se determinó que si se asociaba la prueba de mestizos con la de cruzas dialélicas era posible lograrlo, pues las unidades de selección son líneas S3 , donde la correlación genética entre una línea S3 y su cruza de prueba es 0.93 (Bernardo, 1991), y además la ACG es heredable desde las primeras generaciones de autofecundación de las líneas (Lile y Hallauer 1994).
Cruzas dialélicas entre líneas seleccionadas en mestizos
En el análisis conjunto de las cruzas hubo diferencias significativas (P ≤ 0.05) para todas las variables estudiadas en todos los factores de variación, excepto porcentaje de acame de tallo en cruzas e interacción cruzas × ambientes, resultados que indican un comportamiento diferencial entre ambientes, cruzas y la respuesta de éstas a través de los ambientes de evaluación (Cuadro 5). Con base en estos resultados se realizó el análisis genético de las cruzas, en el que se encontró que los efectos de ACG y ACE fueron significativos (P ≤ 0.05) para todas las variables, excepto los efectos de ACE para floración masculina y porcentaje de acame de tallo, resultados que indican que las líneas evaluadas difieren tanto en sus valores genéticos aditivos como no aditivos (Cuadro 5).
Cuadro 5. Cuadrados medios del rendimiento de grano y características agronómicas de cruzas dialélicas formadas con nueve líneas de la población ST C2 F2 sobresalientes en prueba de mestizos. Análisis combinado a través de cinco localidades. 2016.
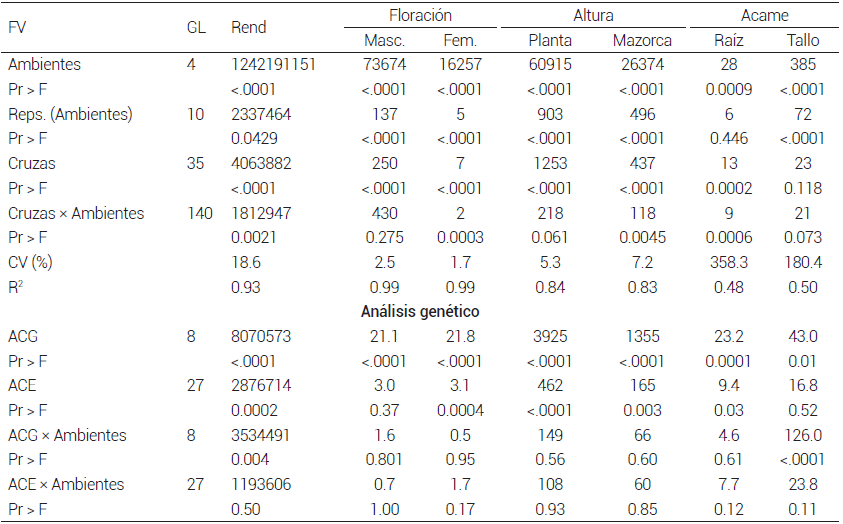
FV:fuente de variación, Rend: rendimiento de grano.
La interacción ACG × ambientes resultó significativa (P ≤ 0.05) para rendimiento de grano y acame de tallo, mientras que en la interacción ACE × ambientes no hubo significancia para ninguna de las variables, resultados que coinciden parcialmente con los obtenidos por Murtadha et al. (2018), quienes en un trabajo similar no obtuvieron diferencias significativas en rendimiento de grano en los dos tipos de interacción señalados y en este estudio la interacción ACG × ambientes fue significativa (P ≤ 0.05), lo que muestra que los efectos aditivos son más sensibles al ambiente que los no aditivos (Cuadro 5).
La relación entre el cuadrado medio de la ACG y la ACE fue mayor que 1.0 en todas las variables estudiadas, lo cual muestra que predominó la varianza genética aditiva sobre la no aditiva y se espera que para estas características haya respuesta en los siguientes ciclos de selección en la población ST C2 F2 , lo que concuerda con lo señalado al respecto por Fasahat et al. (2016) y Murtadha et al. (2016). Los progenitores ST C2 F2 -29-2-1, ST C2 F2 -69-3-1 y ST C2 F2 -21-2-1 tuvieron ACG positiva y significativa (P ≤ 0.05) para rendimiento de grano, y serían los que más contribuirían al rendimiento de grano del sintético; los que menos contribuirían serían los progenitores ST C2 F2 -56- 1-2, ST C2 F2 -89-1-1 y ST C2 F2 -77-2-1, pues su ACG fue negativa y significativa (P ≤ 0.05) (Cuadro 6).
Cuadro 6. Aptitud combinatoria general de progenitores seleccionados de maíz derivados de la población ST C2 F2 evaluados en cruzas dialélicas. 2016.
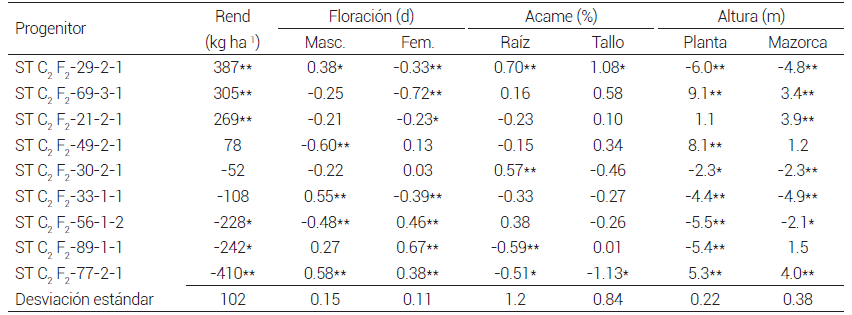
*,**Valores significativos al 0.05 y 0.01 de probabilidad. Rend: Rendimiento de grano.
Al comparar la ACG obtenida para rendimiento de grano en mestizos y cruzas dialélicas, hubo un comportamiento diferencial entre las líneas evaluadas (Cuadros 4 y 6). La línea ST C2 F2 -29-2-1 tuvo ACG alta en las dos pruebas; ésta sería un progenitor ideal para el mejorador porque tiene efectos no aditivos que permitirían obtener buenas combinaciones en cruza con progenitores del otro componente del par heterótico y efectos aditivos altos que contribuirían en el rendimiento de grano de la población; en cambio, la línea ST C2 F2 -56-1-2 tuvo ACG alta y significativa en mestizos (P ≤ 0.05), y negativa y significativa en cruzas dialélicas (P ≤ 0.05), lo que indicaría que predominan los efectos no aditivos sobre los aditivos, ya que en cruza con progenitores del otro componente heterótico se esperarían buenas combinaciones, pero contribuiría poco en el rendimiento de grano de la población.
Un caso contrario al comportamiento de la línea ST C2 F2 -56-1-2 lo constituyeron las líneas ST C2 F2 -21-2- 1 y ST C2 F2 -69-3-1, pues tuvieron valores positivos no significativos de ACG en mestizos y efectos positivos significativos (P ≤ 0.05) en cruzas dialélicas; además, hubo líneas como ST C2 F2 -33-1-1 que tuvieron ACG baja no significativa, tanto en mestizos como en cruzas dialélicas; en este caso, se esperarían combinaciones modestas con líneas del otro par heterótico y baja contribución en el rendimiento de grano de la población (Cuadros 4 y 6).
Al comparar la ACG obtenida para características agronómicas en mestizos y cruzas dialélicas (Cuadros 4 y 6) también se observa que las líneas tuvieron un comportamiento diferencial en ambas pruebas; así, el progenitor ST C2 F2 -29-2-1 mostró consistencia en las dos pruebas en rendimiento de grano, pero no así en acame de raíz y tallo, pues en mestizos tuvo ACG negativa no significativa, pero en cruzas dialélicas tuvo el valor positivo más alto y significativo de ACG (P ≤ 0.05) en acame de raíz y tallo. En este caso el mejorador tendría que decidir si pondera más el rendimiento de grano que la tolerancia al acame. Un caso contrario sería la línea ST C2 F2 -77-2-1, que tuvo la ACG negativa y significativa (P ≤ 0.05) más baja en rendimiento de grano en cruzas dialélicas, pero sus valores de ACG de acame de raíz y tallo fueron negativos y significativos (P ≤ 0.05), por lo que el mejorador tendría que decidir si pondera más al acame que al rendimiento de grano.
Este tipo de ponderaciones son probablemente la razón por la que Moll y Hanson (1984) y Romay et al. (2011) no encontraron respuesta a la selección en SRR en una de las poblaciones del par heterótico, pues muchas veces los mejoradores ceden rendimiento de grano a cambio de tener más tolerancia al acame, y en la SRR con enfoque en hibridación, se pondera más la respuesta a la selección en la cruza que en las poblaciones que la integran; sin embargo, la decisión final dependerá de los objetivos del programa de mejoramiento. En este caso, por ser un programa de hibridación comercial se eliminó al progenitor ST C2 F2 -29-2-1 del sintético por los valores de ACG positivos y significativos (P ≤ 0.05) en acame de raíz y tallo, tomando en cuenta que la tendencia actual es sembrar en altas densidades de población (Egli, 2015).
Con base en la ACG, se considera que el progenitor ST C2 F2 -21-2-1 sería el mejor de este grupo por tener ACG positiva y significativa en rendimiento de grano y valor negativo en acame de raíz, pero los híbridos tendrían plantas con mazorca alta, porque la ACG fue positiva y significativa (P ≤ 0.05) en altura de mazorca (Cuadro 6).
Al analizar la ACE, las cruzas ST C2 F2 -33-1-1 × ST C2 F2 -49-2-1 y ST C2 F2 -21-2-1 × ST C2 F2 -29-2-1 tuvieron la ACE más alta, positiva y significativa, para rendimiento de grano; de éstas dos, la cruza ST C2 F2 -33-1-1 × ST C2 F2 -49-2-1 podría utilizarse como probador para formar cruzas trilineales o dobles utilizando líneas o cruzas élite de la población PABGT-CE C9 F2 , debido a que, además de tener ACE más alta en rendimiento de grano, tuvo valores negativos de ACE en floración masculina y femenina y acame de raíz y tallo, pero se esperaría que las cruzas tengan plantas altas, pues su ACE fue positiva y significativa (P ≤ 0.05) en altura de planta. Otra cruza de interés sería ST C2 F2-30-2-1 × ST C2 F2-89-1-1, pues a pesar de que no fueron significativos los valores de ACG, ésta fue positiva para rendimiento de grano, y negativa para floración masculina y femenina, acame de raíz y tallo, por lo que se esperaría obtener híbridos más precoces y tolerantes al acame, pero con mazorca alta debido a que la ACG de altura de mazorca fue significativa (P ≤ 0.05). Por otro lado, la cruza ST C2 F2 -21-2-1 × ST C2 F2 -29-2-1, que tuvo ACG positiva y significativa (P ≤ 0.05) para rendimiento de grano, se eliminaría debido a que en las cruzas donde la ACE fue positiva y significativa (P ≤ 0.05) para acame de tallo participó la línea ST C2 F2 -29-2-1 (Cuadro 7), lo cual confirma el resultado encontrado en la prueba de ACG (Cuadro 6).
Cuadro 7. Aptitud combinatoria específica entre progenitores seleccionados de maíz derivados de la población ST C2 F2 evaluados en cruzas dialélicas. 2016.
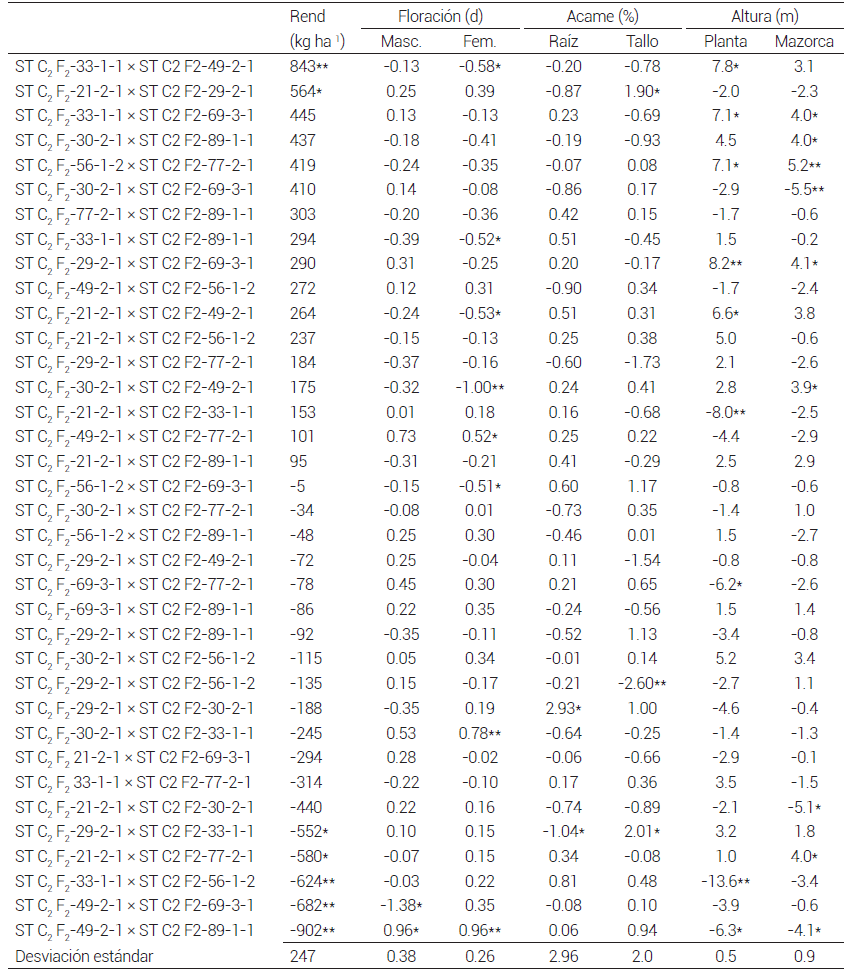
* y **:significancia a P ≤ 0.05 y 0.01, respectivamente. Rend: rendimiento de grano en materia seca (kg ha-1).
En resumen, de acuerdo con los resultados obtenidos, se considera que la prueba de mestizos asociada con la de cruzas dialélicas tiene ventajas importantes debido a que se seleccionan progenitores con valores altos de ACG y ACE, se eliminan progenitores dudosos antes de recombinarse en el siguiente ciclo de selección, se obtiene la F1 y hay mejor recombinación en la F2 entre progenitores seleccionados, además de que se obtienen líneas y cruzas simples para formar híbridos simples, trilineales y dobles con progenitores del otro par heterótico. Las desventajas relativas serían un mayor costo adicional para formar y evaluar las cruzas y que se usa un ciclo agrícola más para completar un ciclo de selección en la SRR.
CONCLUSIONES
Tanto en la prueba de mestizos como en cruzas dialélicas las líneas ST C2 F2 -21-2-1 y ST C2 F2 -29-2-1 sobresalieron por su alta ACG para rendimiento de grano, eliminándose la última porque en cruzas dialélicas la ACG y ACE para acame fue positiva y significativa (P ≤ 0.05). La cruza simple ST C2 F2 -33-1-1 × ST C2 F2 -49-2-1 resultó la más prometedora porque sus valores de ACE fueron positivo significativo (P ≤ 0.05) para rendimiento de grano, negativo y significativo (P ≤ 0.05) para número de días a floración femenina, y negativos, aunque no significativos, para acame de raíz y tallo. Asociar las pruebas de mestizos y cruzas dialélicas permitió seleccionar con más rigor líneas y cruzas con alta ACG y ACE para rendimiento de grano y características agronómicas, eliminar líneas indeseables antes de formar un sintético y obtener líneas y cruzas simples prometedoras para formar híbridos sobresalientes con progenitores élite del otro componente heterótico.

