











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
La mayoría de las plantaciones comerciales en México, provienen de la iniciativa de campesinos con pequeñas propiedades que confían en obtener mejores dividendos de utilidades a los generados por algunos cultivos agrícolas (López-López y Caballero-Deloya 2018). En los últimos años el establecimiento de plantaciones forestales es una alternativa para recuperar terrenos degradados, disminuir las tasas de erosión y asegurar el abastecimiento de madera a la industria (Velázquez-Martínez et al. 2013). Ya que las plantaciones convierten la energía solar en productos útiles, además del secuestro de carbono que es de alrededor de 113 x 106 Mg año-1 y la liberación de aproximadamente 82 x 106 Mg año-1 de O2 (Greaves et al. 2015).
En el estado de Durango se establecen en promedio 3 674 ha año-1 de plantaciones forestales, en donde Pinus durangensis Ehren., Pinus engelmannii Carr., Pinus cooperi Engelm., Pinus arizonica Engelm., Pinus cembroides Zucc, y Pinus chihuahuana Engelm., son las especies más utilizadas, primordialmente en áreas con insuficiente regeneración natural, áreas bajo manejo forestal y con algún grado de alteración en los suelos (Mejía-Bojórquez et al. 2015). Pinus engelmannii Carr. es un pino típico de las Sierra Madre Occidental, se presenta en los estados de Chihuahua, Durango, Sinaloa y Zacatecas (Perry 1991), es una especie maderable de alto valor comercial, muy utilizada en reforestación para programas de conservación y restauración en el noroeste de México (Ávila-Flores et al. 2016).
Las plantaciones forestales adquieren relevancia como sistemas productivos que manejados de manera intensiva y con turnos cortos (20 años) producen mayor cantidad de madera de calidad por unidad de superficie (Romo-Guzmán et al. 2014). Por lo que la industria maderera promueve el establecimiento de plantaciones comerciales que favorezcan el aumento en la proporción de madera juvenil, debido a la intensificación de las prácticas forestales y al impulso de la producción de materia prima rentable (Moore y Cown 2017).
La madera juvenil se extiende a partir de la médula y hasta un intervalo de 14 a 20 anillos de crecimiento, en Pinus patula Schl. Et Cham (Meza-Juárez et al. 2005) y Pinus sylvestris L. (Gryc et al. 2011). Las características y propiedades que presenta la madera juvenil son: anillos de crecimiento anchos, traqueidas cortas, pared celular delgada, grano espiralado y densidad básica clasificada como baja (Larson et al. 2001, Ilic et al. 2003, Jovanovski et al. 2005).
La densidad básica de la madera es una propiedad que permite inferir la calidad de los productos maderables que se pueden ofrecer a la industria (García-Aguilar et al. 2017). Mientras que, el contenido de humedad se relaciona con el transporte de materia prima, el secado de la madera y las etapas posteriores en el procesamiento de una variedad de productos (Moreno-Chan et al. 2012). Las contracciones de la madera afectan directamente la estabilidad dimensional de productos de madera y contribuye a la formación de grietas superficiales e internas, así como alabeos y deformaciones (Liu y Wang 2016).
La densidad básica de la madera también es importante para la conversión del volumen de las existencias reales (m3) a biomasa (t) y en particular para la estimación del contenido de carbono (C), ya que, permite obtener estimaciones precisas de ambos elementos (Ordóñez-Díaz et al. 2015); como lo sustentan investigaciones donde se obtiene la biomasa al multiplicar el volumen por la densidad básica (Barth et al. 2016, Rodríguez-Larramendi et al. 2016, García-García et al. 2019). Hay otras alternativas para la estimación de biomasa como las funciones alométricas basadas en muestreos destructivos que proporcionan estimaciones precisas de la biomasa individual y por unidad de superficie (Soriano-Luna et al. 2015).
En la actualidad, se tiene la necesidad de disponer de información eficiente sobre la cantidad de almacenamiento de biomasa y de los flujos de carbono forestal en los diferentes ecosistemas terrestres, debido a su influencia para mitigar el cambio climático global (Cuevas y Aquino 2020). Por lo que el conocimiento del desempeño de la especie en cuanto a la acumulación de biomasa y sus vinculaciones al crecimiento y calidad de madera obtenida es una herramienta fundamental para la toma de decisiones en la actividad silvícola (Barth et al. 2016).
A pesar de los trabajos realizados sobre Pinus engelmannii Carr., la información es escasa con respecto a las propiedades físicas de la madera proveniente de plantaciones. El objetivo del presente trabajo fue determinar la calidad de la madera de una plantación de P. engelmannii de 17 años mediante la obtención de la densidad básica, contenido de humedad y contracciones volumétricas totales; además de estimar la biomasa de la plantación. La hipótesis formulada es: que existe diferencia en las propiedades físicas entre árboles y dentro de los árboles de la plantación; y existe diferencia entre las ecuaciones utilizadas y la clase diamétrica en el cálculo de biomasa.
Materiales y métodos
El área de estudio se delimitó a una plantación de P. engelmannii, establecida en el Ejido El Encinal, municipio de Durango, México. El clima es templado subhúmedo con lluvias en verano (Cw), con precipitación media anual de 754.6 mm, exposición zenital, topografía plana, suelo profundo con textura fina de tipo Luvisol (INEGI 2018). La plantación se ubica en el polígono con las coordenadas geográficas 23° 42’ 53.1” LN - 105° 03’ 38.7” LO- 23° 42’ 53.1” LN - 105° 03’ 31.0” LO; 23° 42’ 50.4” LN - 105° 03’ 30.4” LO; 23° 42’ 51.4” LN - 105° 03’ 37.9” LO; con altitud de 2 597 m tomados con un GPS eTrex 10 marca Garmin. La superficie plantada fue de 15 ha con planta proveniente de vivero y previo al establecimiento en el año 2002 se realizó la roturación del terreno con maquinaria pesada, el diseño de plantación fue a marco real a la distancia de 1.5 x 1.5 m, dando una densidad de 4 444 árboles ha-1.
El trabajo de campo se realizó en el año 2019, en 20 sitios rectangulares de 3.0 x 6.0 m, a través de muestreo sistemático con separación de 30 m entre sitio sobre la misma hilera y separación de 15 m entre hileras (CONAFOR 2011). A cada árbol presente en el sitio se le tomó la altura total (At) en m con una Pistola Haga (altímetro), el diámetro normal (d) en cm con una cinta diamétrica marca Weiss® modelo 283D de 5 m (Romahn y Ramírez 2010). Para la estimación de la biomasa en el fuste, se consideró un total de 113 árboles, con un intervalo de d entre 8 y 21.01 cm; mientras que la At estuvo entre 5.4 y 13.5 m.
Los cilindros de madera se colectaron de dos árboles dominantes por sitio con At entre 6.5 y 13.5 m, con d entre 12 y 21 cm, sin presencia de daños físicos o ataque de plagas, mediante un taladro de Pressler de 5 mm marca Haglöf®, en 40 árboles a una altura de 1.30 m. Los cilindros de madera, previamente identificados, se colocaron en un recipiente con agua, para mantener la muestra saturada y evitar la pérdida de humedad (Goche-Télles et al. 2011).
Los cilindros de madera de cada árbol en estado saturado se dividieron en cinco secciones a partir de la médula y en dirección de la corteza, en cada sección se midió el largo con un vernier digital marca Mitutoyo®, para el diámetro saturado se consideró un valor de 5 mm (diámetro del taladro); el peso saturado se obtuvo con la balanza analítica marca Velab® con aproximación de 10 milésimas de gramo (ASTM-D2395 2017). Posteriormente, las secciones se colocaron en cajas Petri y se introdujeron a la estufa de secado Grieve® modelo LW -201C, a temperatura de 100 ± 2°C, hasta obtener un peso seco constante (peso anhidro), el cual se logró a las 48 horas (Goche-Télles et al. 2011). A las muestras en estado anhidro se les midió el largo, diámetro y peso (Goche-Télles et al. 2011). Con los datos en estado saturado y anhidro se obtuvo la densidad básica mediante el método de máximo contenido de humedad (Smith 1954).
Dónde: DB densidad básica (g cm-3), Pi peso inicial (saturado) (g), Po peso anhidro (g), 1.53 densidad real de la madera.
El contenido de humedad se obtuvo de acuerdo con la fórmula propuesta por Millers (2013).
Dónde: CH es el contenido de humedad (%), Pi peso inicial (saturado) (g), Po peso anhidro (g).
Las contracciones volumétricas totales se determinaron de acuerdo a la fórmula propuesta por Schonfelder (2018).
Dónde: CVT contracciones volumétricas totales (%), Vv volumen verde (cm3), Volumen anhidro (cm3).
El punto de saturación de la fibra se obtuvo de acuerdo con la fórmula propuesta por Fuentes-Salinas (2000).
Dónde: PSF punto de saturación de la fibra, CVT contracciones volumétricas totales, DB densidad básica.
El volumen se obtuvo con la ecuación presentada por Mejía-Bojórquez et al. (2015) para Pinus engelmannii de Durango.
Dónde: V es volumen en m3, d es el diámetro normal (m), At es la altura total (m) y los dos parámetros estimados. Mientras que la estimación de la biomasa del fuste se hizo mediante la ecuación Y = V * Db * 1000, donde Y es biomasa del fuste (kg), V volumen (m3) y Db densidad básica (g cm-3) (García-García et al. 2019); y tres ecuaciones alométricas, una determinada para P. engelmannii proveniente de rodales naturales del estado de Durango, otra para pinos y una última para P. patula (Tabla 1). Las ecuaciones alométricas utilizaron el modelo básico que relaciona diámetro normal y altura total como variables independientes (Lim et al. 2013). Con lo anterior, se busca fortalecer la importancia de la densidad básica en la determinación de la biomasa. Por otro lado, se clasificaron los árboles en clases diamétricas con una amplitud de 10 cm, encontrando tres categorías 10, 15 y 20 cm de diámetro.
Tabla 1 Ecuaciones alométricas utilizadas en la estimación de la biomasa del fuste (kg) de Pinus engelmannii proveniente de una plantación.
| Nombre | Modelo | Ecuación | Autor (s) |
|---|---|---|---|
| EC-1 |
B=Exp(-β0)*(d2*At)β1
|
B=Exp(-4.682959)*(d2*At)1.033543
|
Soriano-Luna et al. (2015) |
| EC-2 |
B=αdβAtγ+εi
|
B=0.0291*d1.74165*At1.16614
|
Vargas-Larreta et al. (2017) |
| EC-3 |
B=0.09798*d1.67370*At1.02867
|
Ec = Ecuación, B = biomasa del fuste, d = diámetro normal (cm), At = altura total (m), εi = error, y los parámetros estimados α, β.
El análisis de regresión lineal múltiple: Y = β0 + β1X1 + β2X2 + εij se realizó para descubrir si las variables independientes (X1) árbol y (X2) sección dentro del árbol, influyen en las propiedades físicas estudiadas (DB, CH y CVT), consideradas variables dependientes (Y) (Kaya-Uyanik y Nese 2013). El análisis de varianza se determinó con el procedimiento PROC GLM de las siglas en inglés Generalized Linear Models (modelo lineal generalizado) del sistema SAS® (SAS 2002), el cual se utiliza como una alternativa a la transformación de la variable dependiente/respuesta y a la falta de normalidad (McCullagh y Nelder 1989), en las variables que mostraron resultados significativos, se realizaron pruebas de medias de Tukey (α = 0.05). Adicional a lo anterior se efectuaron correlaciones de Pearson para mostrar la relación lineal entre la densidad básica con el contenido de humedad y las contracciones volumétricas totales, para la interpretación del nivel de la magnitud de la correlación se utilizó el cuadro de Cohen (Hernández-Lalinde et al. 2018). El efecto de la clase diamétrica sobre la cantidad de biomasa se analizó mediante un análisis de varianza, y la prueba de medias de Tukey (α = 0.05)
Resultados
Propiedades físicas
La densidad básica, el contenido de humedad y las contracciones volumétricas totales de la madera de Pinus engelmannii en la plantación de 17 años presentaron amplia variación en los valores, como lo reflejan los valores mínimo y máximo reportados para cada una de las propiedades; que es congruente con los valores altos del coeficiente de variación (Tabla 2). Con base en los valores promedio de la densidad básica y las contracciones volumétricas totales se determinó el Punto de Saturación de la Fibra que fue de 39.28%.
Tabla 2 Valores generales de las propiedades físicas estudiadas de Pinus engelmannii.
| Valores | Densidad básica (g cm-3) | Contenido de humedad (%) | Contracciones volumétricas totales (%) |
|---|---|---|---|
| Mínimo | 0.241 | 135.02 | 8.33 |
| Promedio | 0.379 | 199.37 | 13.63 |
| Máximo | 0.499 | 348.80 | 18.64 |
| Desviación estándar | 0.053 | 38.563 | 2.151 |
| Coeficiente de variación (%) | 14.785 | 19.527 | 16.195 |
El análisis de varianza muestra que la sección del árbol influye en la densidad básica y contenido de humedad (P < 0.0001), lo que se confirma con la prueba de comparación de medias de Tukey (α = 0.05), que muestra una diferencia significativa entre las secciones en que se dividió el cilindro de madera, presentando una variación dentro del árbol en sentido radial (médula-corteza) (Tabla 3).
Tabla 3 Variación dentro del árbol de las propiedades físicas de Pinus engelmannii.
| Sección | Densidad Básica (g cm-3) | Contenido de Humedad (%) | Contracciones volumétricas totales (%) |
|---|---|---|---|
| 1 | 0.343 ± 0.051 c | 231.89 ± 40.75 a | 13.66 ± 2.186 a |
| 2 | 0.379 ± 0.055 b | 204.56 ± 47.80 b | 13.46 ± 2.081 a |
| 3 | 0.389 ± 0.044 b | 194.43 ± 25.48 b | 14.14 ± 1.979 a |
| 4 | 0.409 ± 0.059 a | 185.08 ± 47.99 b | 13.66 ± 2.241 a |
| 5 | 0.418 ± 0.053 a | 177.90 ± 37.86 b | 13.21 ± 2.318 a |
Letras iguales por columna indica que no existe diferencia estadística significativa entre secciones (α = 0.05)
La correlación entre la densidad básica y las contracciones volumétricas presentaron una correlación débil (r = -0.14), mientras que con el contenido de humedad presentó una correlación negativa fuerte (r = -0.96, P < 0.0001), (Hernández-Lalinde et al. 2018). mostrando que a medida que la densidad básica aumenta el contenido de humedad saturado disminuye (Figura 1).
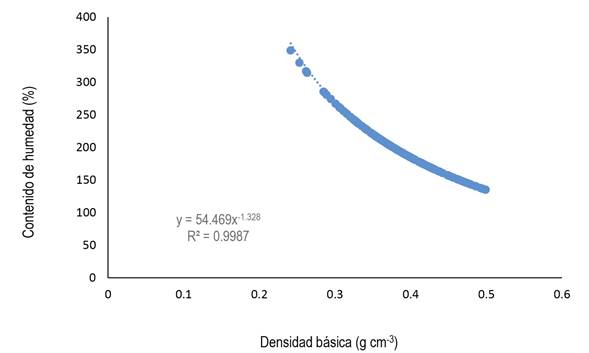
Biomasa fustal
En la plantación cada árbol en promedio tuvo 22.79 kg de biomasa, con las características dasométricas promedio de 13.60 cm y 8.8 m de diámetro normal y altura total, respectivamente. La biomasa real del fuste no presenta diferencias significativas (P < 0.0001) con los estimados por la EC-1 que fue desarrollada para Pinus patula, sin embargo, con las ecuaciones generadas para pinos (EC-2) y Pinus engelmannii (EC-3) si presenta diferencias significativas al estimar la biomasa con los mismos datos (Tabla 4).
Tabla 4 Estadísticos generales de la biomasa estimada del fuste con las diferentes ecuaciones aplicadas.
| Biomasa Real (kg) | Ecuación 1 | Ecuación 2 | Ecuación 3 | |
|---|---|---|---|---|
| Promedio | 22.79 ± 2.14 c | 22.22 ± 2.20 c | 40.06 ± 2.41 b | 80.17 ± 4.65 a |
| Máximo | 100.60 | 97.99 | 141.41 | 276.64 |
| Mínimo | 0.40 | 0.37 | 0.97 | 2.51 |
Letras diferentes en la misma fila indican una diferencia significativa entre las ecuaciones (α = 0.05)
La densidad básica de la madera obtenida por categoría diamétrica de 10, 15 y 20 cm fue de 0.347, 0.383 y 0.396 g cm-3, respectivamente, con estos valores de densidad se calculó la biomasa real del fuste por categoría diamétrica. El promedio de la biomasa presentó diferencias significativas (P < 0.0001) entre las categorías diamétricas presentadas (10, 15 y 20), en cada una de las ecuaciones validadas (Tabla 5). La biomasa presentó la tendencia de incrementar con el aumento del diámetro normal en todas las ecuaciones utilizadas (Figura 2).
Tabla 5 Valores promedio de la biomasa del fuste (kg) encontrado por categoría diamétrica con las ecuaciones estudiadas.
| Categoría diamétrica (cm) | |||
|---|---|---|---|
| 10 | 15 | 20 | |
| Real | 9.82 ± 7.05 c | 25.80 ± 6.15 c | 45.77 ± 18.68 c |
| Ecuación 1 | 12.09 ± 06.69 c | 23.67 ± 6.15 c | 43.47 ± 28.03 c |
| Ecuación 2 | 20.64 ± 10.06 b | 35.20 ± 10.94 b | 57.83 ± 21.31 b |
| Ecuación 3 | 48.95 ± 23.44 a | 86.17 ± 16.18a | 142.26 ± 84.13 a |
Letras diferentes por columna indican diferencias significativas entre las ecuaciones utilizadas (Tukey, α = 0.05)
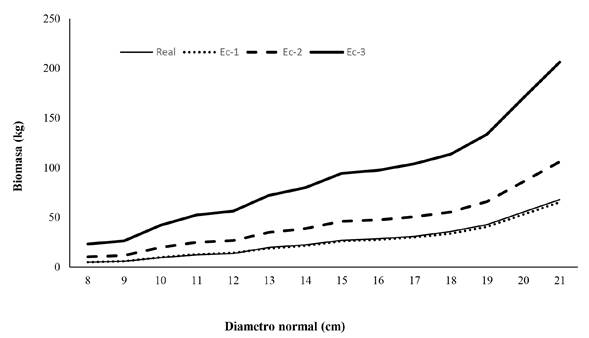
Discusión
Propiedades físicas de la madera de la plantación
El monitoreo de la densidad básica en plantaciones es de suma importancia, puede ser un indicador de la calidad de la madera que se ofrecerá a la industria (García-Aguilar et al. 2017). La densidad básica promedio obtenida en este trabajo fue de 0.379 g cm-3, la cual se clasifica como baja de acuerdo a lo establecido por Sotomayor-Castellanos (2005), el valor fue menor a lo reportado de rodales naturales por Honorato-Salazar y Meraz-Alemán (2002) y Silva-Arredondo y Návar-Cháidez (2012). Se considera que valores menores en la densidad de la madera proveniente de plantaciones respecto a rodales nativos es resultado de un aumento en la proporción de madera temprana por anillo de crecimiento (McLean 2019), así como de madera juvenil (Larson 2001), lo anterior propiciado por una mejor fertilidad del suelo (Watt y Trincado 2019).
En apoyo a lo anterior, se considera que suelos con menor calidad, propician una disminución en el crecimiento de los árboles favoreciendo la producción de madera madura (Sackser et al. 2018) y por consecuencia madera con mayor densidad (Greaves et al. 2015). Esto también fue observado en madera de Pinus elliottii var. elliottii que presentó valores de 0.393 g cm-3 en madera juvenil y 0.479 g cm-3 en madera madura. Esto trae por consecuencia, un retraso en la edad a la que el arbolado alcanza el diámetro mínimo de corta (García-Aguilar et al. 2017), coincidiendo con Reyes-Cuauro et al. (2012), quienes reportan que a 21 años se tienen densidades de 0.504 g cm-3 en Pinus caribaea var. hodurensis.
El contenido de humedad de la madera y su distribución es de interés técnico, su contenido puede variar entre piezas de madera obtenidas de un mismo árbol (Goche-Télles et al. 2000). Valores altos encarecen el transporte, reducen la velocidad y calidad en el secado, así como la facilidad de impregnación, también dificultan su transformación en diversos productos (Moreno-Chan et al. 2012). El CH promedio registrado en la madera de Pinus engelmannii en estado saturado fue de 199.37%, el cual se encuentra en el intervalo de 30 - 200% que presentan los árboles (Rodríguez-Abad et al. 2011). A su vez, Reyes-Cuauro et al. (2012) reportan para Pinus caribaea var. hondurensis de 21 años un C.H. de 133.79%, similar a 134% establecido por Dahlen et al. (2020) en Pinus taeda.
El contenido de humedad cambia con relación a la estación del año en que se obtiene la madera y se reduce a medida que se expone a medio ambiente seco (Visser et al. 2014), por lo que madera recién cortada presenta mayor peso (Ofori y Brentuo 2005). La diferencia en el CH entre árboles se puede atribuir a factores del sitio en donde estos se desarrollan y factores intrínsecos de la especie como características anatómicas, por ejemplo, una fibra con pared celular delgada, su vez presentará mayor proporción de lumen lo que permite mayor captación de agua (Al-Sagheer y Prasad 2010).
Con relación a la contracción volumétrica total, para la madera de los árboles de Pinus engelmannii a la edad de 17 años fue de 13.63% en promedio, esto la clasifica como muy alta según Bárcenas-Pazos (1995). Los resultados coinciden con lo reportado por Riesco-Muñoz y Díaz (2007) en Pinus pinaster (13.8%) y Schonfelder et al. (2018) en madera de Pinus sylvestris L, proveniente de una regeneración de matarrasa (13.1%). En general, estas contracciones ocasionarían la presencia de rajaduras y fuertes deformaciones durante el secado (Vignote y Martínez 2006). En cambio, contracciones menores a las encontrados en este trabajo (12.51%) fueron reportadas por Greaves et al. (2015) en Pinus caribaea (12.9%) y Vega-Britez et al. (2019) en Pinus taeda L. de 25 años.
El punto de saturación de la fibra (PSF) se encuentra entre 28 y 32% para la madera de cualquier especie (Suirezs y Berger 2010). Se define como el contenido de humedad en el momento en el cual la madera ha perdido teóricamente todo el vapor de agua y el agua libre contenida en los lúmenes, y las paredes celulares están saturadas de agua higroscópica (Fuentes-Salinas 2000). Pérdidas de humedad inferiores a éste, indica el inicio de las contracciones de la madera (Sotomayor-Castellanos et al. 2021).
El alto valor (39.28%) del PSF determinado en el presente trabajo, puede ser atribuido a la baja densidad de la madera, ya que a medida que la densidad básica disminuye el PSF aumenta (Sotomayor-Castellanos et al. 2021), lo anterior también fue observado por Riesco-Muñoz y Díaz (2007), cuando Pinus radiata con una densidad de 0.34 g cm-3 presentó un PSF de 51%, en contraste, Fuentes-Salinas (2000) reportó un PSF bajo (29.72%) para una densidad básica de 0.42 g cm-3 en madera de Pinus montezumae Lamb.
Variación radial de las propiedades físicas
La densidad básica de la madera no es homogénea dentro de un mismo árbol, dado que depende de la edad cambial (Riesco-Muñoz et al. 2019). En este trabajo se observó un incremento desde la médula hasta la corteza, lo anterior también fue reportado por Goche-Télles et al. (2011) en Pinus patula de Hidalgo, lo mismo encontró Weber (2005) en Pinus caribaea y Gutiérrez-Vázquez y Flores-Montaño (2019) en Pinus oocarpa. Estas tendencias se pueden atribuir a la existencia de madera juvenil en los anillos cercanos a la médula, ya que estos anillos muestran mayor proporción de madera temprana y menor proporción de pared celular, causadas por un crecimiento rápido (Gryc et al. 2011).
Por otra parte, el contenido de humedad es mayor en la médula y disminuye a medida que se aproxima a la corteza (Tomczak et al. 2021, Missanjo y Matsumura 2016). La mayor capacidad de almacenar agua puede atribuirse a que cerca de la médula existen anillos de crecimiento más anchos, con mayor proporción de madera temprana, paredes celulares más delgadas y lúmenes amplios (Gogoi et al. 2017).
Como se observa en este trabajo, el contenido de humedad está influenciado negativamente por la densidad (r = -0.96), es decir, a mayor densidad básica, se presenta menor humedad tal y como lo reafirma Rosales-Solórzano (2019) con una correlación (r = -0.98, P < 0.05) en su trabajo. En consecuencia, se puede considerar a la densidad básica como el factor principal que influye en la variabilidad del contenido de humedad de la madera (Millers 2013).
Contrario a las tendencias presentadas en el contenido de humedad y densidad, los resultados señalan que la contracción volumétrica total de Pinus engeimannii no presenta una relación definida con la densidad básica en dirección radial (r = -0.14), coincidiendo totalmente con lo establecido por Zeidler (2013)). Diferente tendencia fue reportada por Goche-Télles et al. (2000)) en Pinus ayacahuite var. veitchi, mientras que en Pinus syivestrisSchonfelder et al. (2018) y Riesco-Muñoz et al. (2008) encontraron valores de r = 0.59 y r = 0.62, respectivamente.
Biomasa del fuste
Los resultados de biomasa no presentaron diferencias estadísticas significativas al emplear la fórmula que utiliza la densidad básica y el volumen (Rodríguez-Larramendi et al. 2016, García-García et al. 2019, Rodríguez-Laguna et al. 2021) con la ecuación alométrica que relaciona el diámetro normal y la altura total (Soriano-Luna et al. 2015) (Tabla 5). Lo anterior alienta el uso de la densidad básica como variable fundamental en la estimación de la biomasa (Silva-Arredondo y Navar-Chaidez 2010). En esta investigación la biomasa de los árboles producidos en la plantación fue en promedio 26.11 t ha-1, valor mayor a 10.78 t ha-1 encontrado en una regeneración de 15 años (Chávez-Pascual et al. 2017), y a los 7.5 t ha-1 reportados también en una regeneración de Abies reiigiosa (Razo-Zárate et al. 2013).
Por lo que se pudo observar, la biomasa del fuste incrementa con el aumento del diámetro normal, coincidiendo totalmente con Razo-Zárate et al. (2013)), Boca et al. (2017) y Hernández-Moreno et al. (2020), quienes reportan incrementos en densidad básica en sentido radial (Tabla 4), corroborando la estrecha relación de la densidad básica con la biomasa (Rodríguez-Laguna et al. 2021). Otros factores que infieren en la biomasa acumulada son las dimensiones de los árboles (Chávez-Pascual et al. 2017), así como la edad del arbolado (Rodríguez-Larramendi et al. 2016). La misma tendencia se presentó con las cuatro ecuaciones aplicadas (Figura 2), por el contrario, Magalhães et al. (2021) en Pinus ellioti y Pinus taeda, no encuentran diferencias entre las categorías diamétricas. Al respecto, Chávez-Pascual et al. (2017) manifiestan que la calidad del sitio, la densidad de plantación y las dimensiones de los árboles determinan la intensidad de acumulación y cantidad de biomasa aérea.
Conclusiones
La madera de Pinus egnelmannii producida se clasifica como una madera de baja calidad al considerar la densidad básica, la contracción volumétrica alta la hace susceptible a rajaduras y de formaciones. El alto contenido de humedad la hace una madera pesada al momento de transportarla del bosque a la industria. Las características físicas de la madera como densidad básica, el contenido de humedad y las contracciones volumétricas son diferentes entre árboles y aún dentro del mismo árbol. El cálculo de la biomasa se puede realizar mediante ecuaciones que utilizan el volumen y la densidad básica de la madera, debido a que realizan estimaciones similares a las obtenidas por ecuaciones alométricas. La cantidad de biomasa incrementa con el aumento del diámetro normal, y la densidad básica, lo que fortalece la estrecha relación que tiene la biomasa con la densidad básica de la madera.

