











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El concepto de selva inundable (SI) hace referencia a la vegetación que se desarrolla sobre suelos anegados temporal o permanentemente, donde la composición de especies y la estructura de las comunidades varía con la frecuencia y los ciclos de inundación (Palacio-Aponte et al. 2002). Lo habitual es encontrar este tipo de vegetación en los bordes u orillas de algún cuerpo de agua que al desbordarse o aumentar su cauce inunda las áreas en las que se desarrolla (Marks et al. 2014). La SI se desarrolla en planicies que se inundan en alguna época del año, debido a la acumulación del agua de lluvia y permeabilidad del suelo, conociéndose en el sureste de México como vegetación de bajos o Ak'alches (Tun-Dzul et al. 2008, Villalobos-Zapata y Mendoza-Vega 2010). La SI ubicada en el municipio de Calakmul, es una matriz con condiciones ambientales peculiares con predominio de selva subperennifolia (Galindo-Leal 1999, Palacio-Aponte et al. 2002).
La composición y estructura de la vegetación en el municipio de Calakmul se relaciona con los gradientes topográficos, la dinámica de nutrientes, las características del suelo, la geología y las actividades humanas (Martínez y Galindo-Leal 2002). La SI se encuentra situada en planicies acumulativas rodeada de lomeríos en forma de cuenca, los procesos aluviales de sedimentación permiten la acumulación de materiales orgánicos e inorgánicos que inhiben la infiltración y permiten la acumulación de agua en el suelo (Palacio-Aponte et al. 2002). Este ecosistema es fuente de agua para los habitantes de las localidades, hábitat de especies epífitas y refugio o área de anidación y reproducción de la fauna (Villalobos-Zapata y Mendoza-Vega 2010). Desafortunadamente, como el resto de las selvas de la región, la SI está inmersa en procesos de cambio de uso de suelo que originan deforestación y fragmentación (García-Licona et al. 2014). Al respecto, Martínez y Galindo-Leal (2002) mencionan que la SI de Calakmul ha sido objeto de explotación selectiva, lo que posiblemente favoreció cambios en la composición de especies. A pesar de su importancia ecológica y de los problemas que enfrenta la SI en este municipio, se tienen pocos estudios que describan su diversidad y estructura florística (Tun-Dzul et al. 2008). Por lo anterior el objetivo del trabajo fue describir la diversidad y la estructura de especies arbóreas en la SI al sur y centro del municipio de Calakmul.
Materiales y métodos
Área de estudio
El estudio se realizó en nueve localidades del municipio de Calakmul, que se encuentran ubicadas entre los 19° 12' 00" y 17° 48' 39" LN y, 89° 09' 04" y 90° 28' 05" de LO (Figura 1). Las cuales fueron: Centauro del Norte (CN), El Carmen II (EC), Narciso Mendoza (NM) y Nuevo Conhuás (NC) al centro del municipio; Arroyo Negro (AN), Carlos A. Madrazo (CM), Josefa Ortiz Domínguez (JOD), Ley de Fomento Agropecuario (LFA) y Unidad y Trabajo (UT) al sur del municipio. Los sitios se caracterizan por tener más de 40 años sin usarse para actividades agropecuarias o forestales, sin evidencias de actividad humana reciente con selva baja subperennifolia, suelo tipo Ak'alche. Posiblemente, al igual que el resto de las selvas de la región de Calakmul, hayan estado expuestas a extracción de madera y de chicle entre 1930 y 1970 (Kepleis 2004). El clima del municipio es cálido subhúmedo (García 1988). La temporada de lluvias comprende de junio a noviembre con precipitación media anual de 1 076.2 mm, y temperatura promedio de 24.6 °C (Martínez y Galindo-Leal 2002). La altitud en el centro del municipio es de 200 a 250 msnm con declives menores al 10 %, al sur pueden encontrarse altitudes menores a los 100 msnm (INEGI 2014).
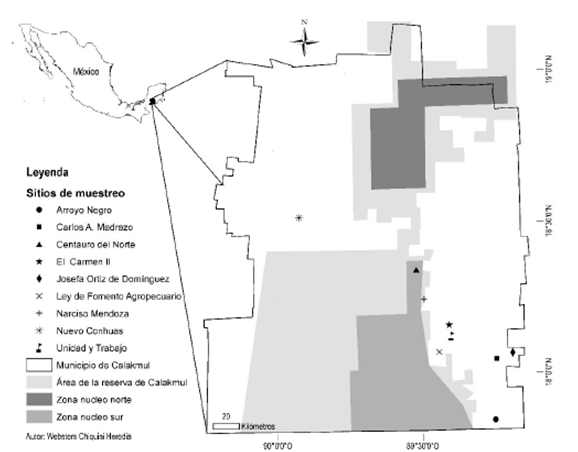
Diseño de muestreo
La selección de las comunidades para la instalación de las parcelas se realizó al azar, tomando en cuenta todas las comunidades al centro y sur de Calakmul en las cuales se tiene registró de la SI, mientras que la selección de parcelas en cada comunidad se realizó de forma sistemática. Se establecieron sitios de muestreo circulares de 1 000 m2 de 17.84 m de radio, con un círculo concéntrico de 400 m2 de 11.28 m de radio. En los círculos de 400 m2 se censaron todos los individuos que a 1.30 m del suelo tuvieran un diámetro normal (Dn) igual o mayor a 2.50 cm y a partir de los 10 cm en el resto del círculo de 1 000 m2. Los individuos que cumplieron con los criterios se etiquetaron, registró la altura total (m), el Dn (cm), y su clasificación taxonómica de acuerdo con las claves respectivas.
Análisis de la información
Con la información de identidad taxonómica de campo, se generó un listado florístico con el orden, familia, género y especie. Se estimó la riqueza de especies, como el número de especies por unidad de muestreo. La diversidad se calculó con el índice de Shannon-Wiener (H’), que permite conocer como varía la riqueza y la distribución de los individuos entre las especies. A partir de los valores de equitatividad (J’) se comparó que tan bien se ajusta la diversidad observada con la diversidad máxima esperada, las cuales se calcularon con las fórmulas de Begon et al. (2006) y Magurran (2004):
Donde: H = índice de Shannon-Wiener, Pi = Abundancia relativa, Log = Logaritmo base 10.
Donde: H'MAX= ln S, S= Número de especies y H'= Índice de Shannon-Wiener.
Para determinar diferencias en la diversidad de los sitios se realizaron verificaciones por pares entre los nueve sitios con la prueba de t-student (Hutcheson 1970). Para estimar la similitud se empleó un análisis de agrupamiento con la medida de Bray-Curtis, para visualizar de forma gráfica la distribución de las parcelas de acuerdo a la similitud que comparten (Calderón-Mandujano et al. 2008). Los cálculos se realizaron con el programa Past® versión 2.17c (Hammer et al. 2001). También se estimó la densidad como el número de individuos por unidad de área (individuos hectárea-1), el área basal (Dn/2)2 π. El valor de importancia relativa (VIR) revela la importancia ecológica de cada especie en una comunidad vegetal, que utiliza parámetros estructurales básicos como abundancia relativa (AR) (individuos ha-1), frecuencia relativa (FR) y dominancia relativa (DR) (m2 ha-1) (Magurran 2004):
Donde:
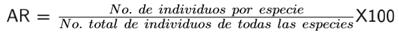
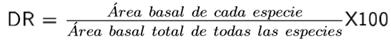
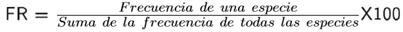
El análisis de estructura horizontal (Dn) y vertical (altura) de la vegetación se realizó con las distribuciones de frecuencia agrupadas por clase, empleando la fórmula K=1+3.33 x log(n), donde K se refiere al número de clases y n el número de árboles. La diversidad estructural se estimó de la diversidad de especies, la altura y el área basal (Martínez-Sánchez 2016). Los indicadores evaluados fueron el índice de Shannon-Wiener para las especies (Hs), el índice de Shannon-Wiener para clases diamétricas (Hd), el índice de Shannon-Wiener para clases de altura (Hh) y el índice promedio de la diversidad estructural (Hsdh), con las fórmulas:
Donde pi= proporción del área basal para la especie i, y m= número de especies
Donde pi= proporción del área basal por clase la diamétrica i, y d= número de clases diamétricas
Donde pi= proporción del área basal para la clase de altura i, y h= número de clases de altura
Donde Hs= diversidad por especies, Hd= diversidad por clases diamétricas, y Hh= diversidad por clases de altura. Para determinar diferencias significativas en la diversidad estructural de los sitios se realizaron verificaciones por pares entre los nueve sitios con la prueba de t-student (Hutcheson 1970).
Resultados
Composición de la comunidad
Se midieron 2 214 árboles, que pertenecen a 112 especies, 84 géneros, 36 familias y 21 órdenes en un área de 0.9 ha (Tabla 1). Las familias mejor representadas fueron Fabaceae (29 especies), Rubiaceae (10), Sapotaceae (7), Apocynaceae (6) y Polygonaceae (6). Estas cinco familias representan el 47 % del total de especies registradas, el resto de las familias están constituidas por cinco o menos especies. De las 112 especies solamente tres (2.68 %) se comparten en los nueve sitios, las cuales fueron Hampea trilobata, M. brownei y Vitex gaumeri; y 32 especies (28.57 %) fueron especies únicas. De las especies únicas, NM contabilizó ocho especies (12.50 %) de la riqueza del sitio, mientras que AN (25 %) y EC (15 %) registraron seis especies, y LFA cinco especies (8.93 %). En el resto de los sitios se contabilizaron menos de tres especies, siendo UT el único sitio que no presentó especies únicas.
Tabla 1. Listado de las especies arbóreas y sus abundancias por sitio de 1 000 m2.

1: Arroyo Negro; 2: Carlos A. Madrazo; 3: Centauro del Norte; 4: El Carmen II; 5: Josefa Ortiz de Domínguez; 6: Ley de Fomento Agropecuario; 7: Narciso Mendoza; 8: Nuevo Conhuás; 9: Unidad y Trabajo. G: gremios; P: Pionera, I: Intermedia, S: tolerante a la sombra.
Diversidad de especies y equitatividad
Los sitios que tuvieron mayor riqueza fueron LFA y NM con 56 especies, seguido de CN con 51 especies (45.53 %), mientras que AN registró la menor riqueza con 24 especies (21.43 %). Los mayores valores de diversidad se obtuvieron en CN (1.558), LFA (1.537) y NM (1.503); mientras que AN (1.057) obtuvo el valor más bajo de diversidad. Los valores de equitatividad muestran que CN presentó la mejor distribución de abundancia entre las especies al ajustarse más de 0.9 la diversidad observada con la diversidad máxima esperada.
Mientras que AN y CAM son los sitios que tuvieron los menores valores de equitatividad, lo que indica mayor abundancia de alguna especie, por los valores menores a 0.8 (Tabla 2). Los sitios muestran diferencias significativas (p > 0.05) en sus valores de diversidad, con excepción de CAM con EC (t= 1.51) y JOD (t= 0.44); CN con LFA (t= 0.62) y NM (t= 1.80); EC con JOD (t= 1.78) y NC (t= 0.92); así como LFA y NM (t= 1.12), NM con UT (t= 1.81).
Similitud en composición de especies
El análisis de agrupamiento (r= 0.8812) mostró que la SI localizada en AN es diferente de las otras SI, lo que indica que la composición de especies es diferente (Figura 2). Además, se observa que las SI con mayor similitud (mayor al 50 %) son las que se localizan en CN, LFA, NC y NM. El sitio CN comparte 37 especies, 72.55 % de la riqueza del sitio con el sitio LFA (66.07 %), 32 especies con NC (80 %) y 35 especies con NM (62.5 %). En tanto que el sitio LFA comparte 32 especies, 57.14 % de las especies con NC (80 %) y 37 especies con NM (66.07 %). Mientras que el sitio NC comparte 31 especies, 77.5 % de la riqueza en el sitio con NM (55.36 %). Por otro lado, las SI de CAM, EC y UT forman un grupo, a un nivel de similitud menor al 40 %.

Estructura de la vegetación
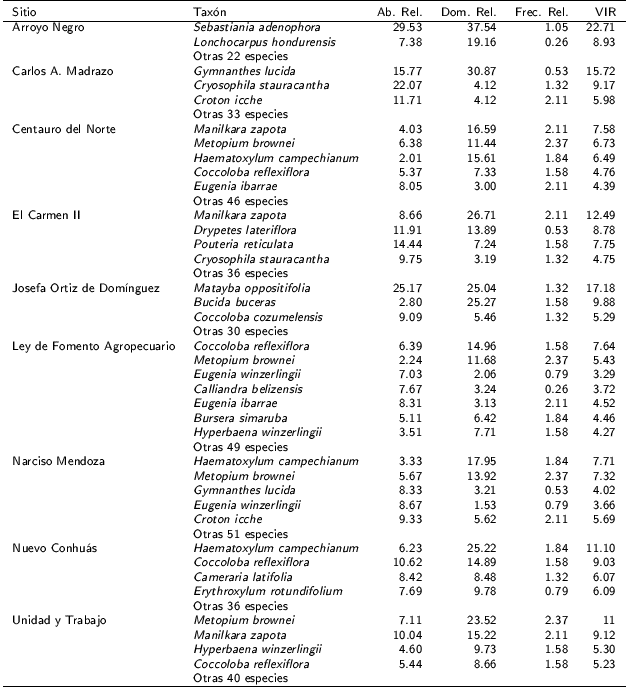
La mayor densidad de individuos se presentó en las SI de LFA, seguida de NM y CN, mientras los valores más bajos se presentaron en JOD y AN. Los valores más altos de área basal se presentaron en las SI de CAM, JOD, UT y NC, mientras que en AN se estimaron las menores áreas básales. Por otro lado, las especies con mayores VIR fueron S. adenophora en AN, M. zapota en CN y EC, M. oppositifolia en JOD, C. reflexiflora en LFA, M. brownei en UT, G. lucida en CM y H. campechianum en NC y NM (Tabla 3).
Tabla 3. Lista de las especies que alcanzaron mayor VIR en cada sitio, el corte se realizó al alcanzar el 30 %.
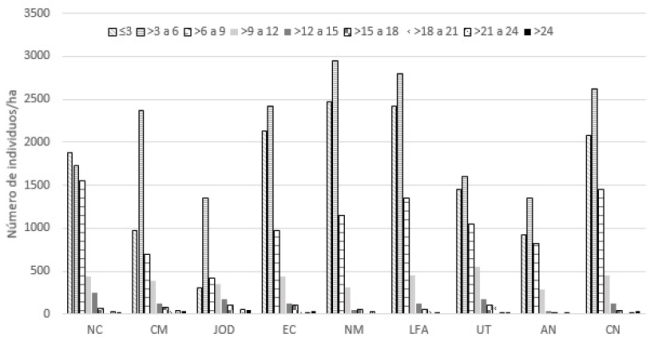
El análisis de la estructura horizontal mostró que, en todas las unidades de muestreo, la mayor proporción de individuos (más del 70 %) se agrupó dentro de las tres primeras clases diamétricas 3, >3 a 6 y 6 a 9 cm, en todos los sitios. Las curvas presentan la forma característica de J-invertida, es decir que hay una baja proporción de individuos con Dn grandes y una alta proporción de individuos pequeños, lo que indica el potencial de regeneración de la vegetación a partir del reclutamiento de individuos (Figura 3).
La altura de los individuos estuvo en el rango de 3 a 13 m, con más del 80 % de los individuos en la tercera y sexta categoría, con distribución en forma de campana (Figura 4).

Diversidad estructural
Al comparar los índices de diversidad estructural Hs, Hd, Hh y Hsdh, se encontró que AN presenta en todos los casos los valores de diversidad estructural bajos (Tabla 4). En el caso de Hs, AN presenta valores significativamente menores (p > 0.001) que el resto de los sitios analizados, mientras que NM y LFA presentan un valor significativamente mayor (p > 0.001) que UT. Para los valores de Hd, no hay diferencias significativas entre las SI. Los valores de Hh muestran que JOD tiene un valor significativamente mayor (p > 0.001) que CM, AN y CN. En el caso de la Hsdh los valores más altos se presentaron en NM y LFA, teniendo los valores más bajos en AN y CM.

Discusión
Composición de la comunidad
Las familias con mayor número de especies coinciden con los reportes para selvas subperennifolias de la península de Yucatán (Zamora-Crescencio et al. 2008, Zamora-Crescencio et al. 2012, García-Licona et al. 2014) y otros lugares (Damasceno-Junior et al. 2005). El número de especies es mayor que el reportado para otras selvas inundables de México (Cortés-Castelán e Islebe 2005, Zamora-Crescencio et al. 2008, Maldonado-Sánchez et al. 2016). La familia Fabaceae es importante por su alto número de especies (Martínez y Galindo-Leal 2002), en este trabajo se registró mayor número de especies de las que obtuvo Zamora-Crescencio et al. (2008) y Maldonado-Sánchez et al. (2016) en selvas del sur de México, pero el número de especies es menor que las reportadas por González-Valdivia et al. (2012). Las diferencias antes mencionadas pueden deberse a que las especies de la familia Fabaceae son indicadoras de vegetación secundaria y las selvas inundables estudiadas es muy probable que hayan estado expuestas a la extracción de madera y chicle (Martínez y Galindo Leal 2002, Kepleis 2004). Las 112 especies que se identificaron, tienen su distribución natural dentro de los límites de la república mexicana (CONABIO 2015), de las cuales C. stauracantha y Handroanthus chrysanthus se encuentran dentro de la categoría de riesgo, bajo el estatus Amenazada en la Norma Oficial Mexicana 059 (SEMARNAT 2010). Las especies Ateleia gummifera y V. gaumeri se encuentran en la categoría de riesgo bajo en el estatus de en peligro de extinción de la Unión Internacional para la Conservación de la Naturaleza (IUCN 2015). Mientras que la especie Swietenia macrophylla se encuentra en la Categoría II de la Convención sobre el Comercio Internacional de Especies Amenazadas de Fauna y Flora Silvestres (CITES 2015). Estas especies son de interés nacional e internacional por su status de conservación o control de su explotación, por lo que pueden tomarse como base para generar estrategias de manejo y conservación en las áreas de selva inundable. De las 112 especies se determinó el gremio al que pertenecen 80 especies, de los cuales 11 son pioneras (9.82 %), 60 intermedias (53.57 %) y 9 tolerantes a la sombra (8.04 %), en los nueve sitios estudiados hay presencia de los tres tipos de especies (Tabla 1). Las especies Haematoxylum campechianum, Cameraria latifolia, Tabebuia rosea, Erythroxylum rotundifolium, Myrciaria floribunda y Ficus cotinifolia, presentes en las SI estudiadas, se han reportado como especies características de selvas inundables por Martínez y Galindo Leal (2002), Vázquez Negrín et al. (2010), Maldonado-Sánchez et al. (2016).
Diversidad de especies y equitatividad
La mayor riqueza registrada por sitio, fue de 56 especies en LFA y NM, que es menor a las 65 especies reportadas por Díaz-Gallegos et al. (2002), para el ejido La Guadalupe en un área de 0.1 ha, pero supera a las 22 especies reportadas por Vázquez-Negrín et al. (2010); lo que puede deberse a la diferencia en los criterios utilizados en la medición de los individuos. El sitio de muestreo del ejido NC registró una riqueza de 40 especies en una superficie de 0.1 ha, similar a la riqueza reportada por Tun-Dzul et al. (2008) de 39 especies en una superficie de 0.05 ha en la misma comunidad. Aunque la cantidad de especies que se comparten es superior al 50 % para la mayoría de los sitios, la abundancia de las especies influye en los resultados de las pruebas estadísticas al comparar la diversidad. La menor diversidad que presentan los sitios al sur del municipio podría deberse a la mayor precipitación (Martínez y Galindo-Leal 2002) que junto con las características topográficas del sitio y de las áreas que la circundan influyen en la inundación (Palacio-Aponte et al. 2002, Cortés-Castelán e Islebe 2005). En los sitios AN, JOD y CM al sur del municipio, se presentaron especies con una alta dominancia, como S. adenophora, Matayba oppositifolia y Chryosophila stauracantha respectivamente. La dominancia de estas especies podría estar relacionado con lo que indican Martínez y Galindo Leal (2002) sobre la influencia que tienen las diferencias en los rangos de humedad al sur del municipio, lo que permite a las especies con mejor capacidad de adaptación crear comunidades dominantes.
Similitud en composición de especies
Las especies H. campechianum, Cameraria latifolia, Bucida buceras y M. brownei se encuentran distribuidas entre los sitios y son indicadoras de terrenos inundados en la región de Calakmul (Palacio-Aponte et al. 2002). Las mayores similitudes se obtuvieron en los sitios que se ubican al centro del municipio. Al respecto Martínez y Galindo-Leal (2002) indican que en el centro del municipio de Calakmul, se tienen condiciones homogéneas de humedad y topografía, lo que permite que se tengan condiciones para el establecimiento de un mayor número de especies. En los sitios al sur se tienen las menores similitudes sobresaliendo los sitios AN y JOD, con una composición de especies diferente, en los cuales no está presente H. campechianum, lo que indica que probablemente estos sitios tienen mayores profundidades de inundación o corrientes superficiales. En AN no se registraron especies de la familia Sapotaceae, probablemente por la composición del suelo (Cortés Castelán e Islebe 2005) o por la poca disponibilidad de oxigeno (López 2009). Al respecto Martínez y Galindo-Leal (2002) mencionan que las diferencias en la abundancia de especies tienen importancia ecológica al permitir que una amplia variedad de fauna pueda hacer uso de estas extensiones de SI, y coincide con Palacio-Aponte et al. (2002) quienes plantean el potencial que tiene la SI de Calakmul como humedal terrestre.
Estructura de la vegetación
Los valores de las áreas básales en los sitios son menores a lo reportados por Maldonado-Sánchez et al. (2016) para selvas inundables de Tabasco, cercanos a lo que reportan Cortés Castelán e Islebe (2005) para selvas inundables de Quintana Roo y Zamora-Crescencio et al. (2008) para Calakmul. Las especies H. campechianum, M. zapota y M. brownei tienen los primeros lugares de VIR por los valores de dominancia que tuvieron, lo que indica que son individuos grandes. Estas especies también fueron reportadas como importantes por Díaz-Gallegos et al. (2002) y Palacio-Aponte et al. (2002). Las especies G. lucida, M. oppositifolia y S. adenophora, no se reportan en otros trabajos, a pesar de que ocupan los primeros lugares de VIR en este estudio para los sitios del sur de Calakmul. La vegetación no presentó diferencias en los patrones de la estructura horizontal y vertical entre los sitios del centro y sur, los individuos más grandes están representados en las clases pequeñas, lo que indica una buena condición de los sitios estudiados.
Diversidad estructural
En general, los sitios NM, JOD y LFA fueron los más diversos estructuralmente, mientras que el sitio AN fue el de menor diversidad estructural. La diversidad estructural asociada con las clases diamétricas (Hd) no tuvo diferencias significativas entre los sitios, al contrario de las clases de alturas (Hh), lo que puede estar asociado a que el atributo de altura puede estar influenciado por la inundación del sitio, pues se sabe que modifica las propiedades del suelo reduciendo la disponibilidad de oxígeno, lo que impacta en el crecimiento de los individuos y la capacidad del suelo para soportar individuos grandes (López 2009).
Conclusiones
El tipo de vegetación en los sitios es de selva baja subperennifolia inundable. Las especies indicadoras de este tipo de vegetación como H. campechianum y C. latifolia, entre otras fueron reportadas para este trabajo, detectando especies con altos valores de importancia que no han sido reportados para la selva de Calakmul, como G. lucida, M. oppositifolia y S. adenophora. La diversidad estructural indica que la altura está influenciada por las inundaciones. La selva inundable de Calakmul tiene potencial para desarrollar programas de conservación, debido a que cuenta con especies prioritarias de atención en los programas nacionales e internacionales y a la importancia que tiene como aporte de agua y alimento a la fauna de la región.

