











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkLas selvas altas perennifolias representan uno de los ecosistemas más vulnerables en el mundo, al poseer los más elevados valores de riqueza de especies que cualquier otro ecosistema terrestre aunado a un fuerte deterioro debido a las actividades del ser humano (Challenger & Soberón 2008, Hernández-Ruedas et al. 2014, Clark et al. 2015, Durán-Fernández et al. 2016). Según Leopold (1950), el área potencial de lo que denominó “tropical rain forest” era alrededor de 12.8 % de la superficie del territorio mexicano. Posteriormente, Rzedowski (1978, 1998) estimó dicha superficie en 11 %, refiriéndose al “bosque tropical perennifolio”; sin embargo, este autor señala: “de tal área, en la actualidad sólo la décima parte ostenta tal vez una vegetación boscosa que por sus características florísticas se asemeje al clímax”. Una estimación más específica y precisa para la selva alta perennifolia posiblemente sea la de Flores-Mata et al. (1971): 74,562 km2 (3.78 % de la superficie del país). Actualmente, la tendencia real es la disminución del área selvática, y probablemente más de 90 % de la extensión que originalmente cubría esté ocupada principalmente por terrenos agrícolas, praderas y comunidades de vegetación secundaria (Rzedowski 1978). Esta tendencia de cambio ha sido ratificada por Challenger y Soberón (2008) al reconocer que la distribución original de las selvas húmedas (que incluye selvas altas, medianas perennifolias y subperennifolias) abarcaba cerca de 9.1 % del territorio nacional (unos 17.82 millones de ha) y se ha reducido en 2002 a 4.82 % (3.16 millones de ha de vegetación primaria, madura o bien conservada, y 6.31 millones de vegetación secundaria). Así, la tendencia de trasformación y remoción es aún más drástica si se considera que para 2005 la superficie sólo de la selva alta perennifolia era de apenas 14,184 km2 (0.72 % de la superficie del país (Challenger y Soberón 2008).
La región Lacandona preserva algunos de los remanentes más extensos de selvas altas perennifolias y mejor conservados en México y es refugio de un número importante de especies de flora y fauna (Martínez et al. 1994, Levy-Tacher et al. 2006, Van Breugel et al. 2006, Avila-Torresagatón et al. 2012, Garmendia et al. 2013, Hernández-Ruedas et al. 2014, Durán-Fernández et al. 2016). Es así que ha sido considerada como una de las áreas de mayor prioridad para la conservación de la biodiversidad en el país, a pesar de que en menos de cinco décadas su cobertura selvática ha disminuido en más del 50 %. Como consecuencia de las actividades humanas en las regiones tropicales del mundo, se aprecia una clara tendencia continua y creciente hacia la eliminación y modificación de las selvas húmedas, principalmente por: (i) aumento de la población humana y su correspondiente demanda de alimentos y otros recursos; (ii) extracciones desmesuradas de los recursos selváticos provocando fuertes procesos degradativos; (iii) orientaciones equívocas al considerar inagotables los recursos de la selva; y (iv) poca atención a su conservación y al desarrollo de sistemas de aprovechamiento persistente (Miranda 1975, Farnworth & Golley 1977, Nelson 1977, Muench-Navarro 1978, Rzedowski 1978, UNESCO 1980, Nations & Nigh 1980, Casco-Montoya 1984, Anderson 1990, Vásquez-Sánchez & Ramos-Olmos 1992, FAO 1994, Martínez et al. 1994, Challenger & Soberón 2008, Hernández-Ruedas et al. 2014).
Los cambios de la acción humana sobre los ambientes tropicales han causado no sólo la destrucción gradual del recurso y su potencial económico, sino también la pérdida de los conocimientos tradicionales sobre su manejo, junto con valores éticos y culturales (Schultes 1991, Phillips & Gentry 1993). Así, para el caso de Mesoamérica, Mac Neish (1964), Wilken (1971) y Barrera et al. (1977) han resaltado la importancia del manejo y aprovechamiento de la selva para el desarrollo de la civilización maya. Levy-Tacher & Golicher (2004) y Levy-Tacher et al., (2002, 2005) han acreditado la importancia del cocimiento tradicional lacandón sobre el medio ecológico y manejo silvícola que debe ser considerado en los estudios sobre flora y vegetación de la región. Además, cabe resaltar que los agricultores de estas regiones dependen directamente de la persistencia de estos recursos para su propia subsistencia y sobrevivencia.
La tendencia actual en el uso del suelo en la Selva Lacandona es similar a la de otras regiones tropicales del mundo. Así, uno de los problemas más serios que enfrenta la región Lacandona, es la disminución de su área selvática, como consecuencia de un fuerte proceso de colonización, la expansión de las vías de comunicación, la tala inmoderada de las masas forestales y la incorporación de nuevas áreas de selva al aprovechamiento agrícola y pecuario (Rzedowski 1978, Nations & Nigh 1980, Muench-Navarro 1982). Por ello, el estudio de las comunidades maduras de selva y el manejo que reciben por parte de los agricultores autóctonos, es importante antes de que desaparezcan y con ello se pierda la oportunidad de registrar su flora y fauna y el conocimiento tradicional respectivo sobre su biología y utilidad (Levy-Tacher et al. 2002, 2005, 2006).
El objetivo del presente estudio fue describir de forma detallada la estructura de la vegetación madura de la selva alta perennifolia en la comunidad Lacandona de Nahá, su variación estructural, así como la composición florística y sus formas vitales, con el apoyo de informantes lacandones locales, quienes cuentan con un vasto conocimiento de la vegetación natural de su región. Esta información es de suma importancia como referencia ante los efectos de los aprovechamientos, de las prácticas de rehabilitación en áreas deterioradas, y para reconocer oportunamente la necesidad de aplicar medidas correctivas de aprovechamiento con fines conservacionistas.
Materiales y métodos
Zona de estudio. El estudio se realizó en los bienes de uso comunal dentro de la comunidad Lacandona de Nahá, en el municipio de Ocosingo, Chiapas (Figura 1), la cual fue decretada como Área Natural Protegida en 1998, con una superficie de 3,847 ha, localizada entre los paralelos 16º 56’ y 17º 00’ de latitud norte y los meridianos 91º 32’ y 91º 37’ de longitud oeste (INEGI 1988a, SEMARNAP-SRA 1998, SEMARNAT-CONANP 2006). Müllerried (1957) incluyó esta porción de la región Lacandona dentro de lo que denominó región natural de montañas marginales del norte y oriente de Chiapas. Su fisiografía está formada por serranías bajas, con elevaciones que alcanzan hasta 1,280 m s.n.m., alternadas con valles, geoformas que Muench-Navarro (1982) consideró como serranías y terrazas antiguas. Geológicamente predominan las calizas del cretácico superior (Müllerried 1957, INEGI 1988b). El sistema fluvial superficial se manifiesta en una exigua red de ríos y arroyos que desembocan en los ríos Tulijá y Santo Domingo-Chocoljá, los cuales drenan en dirección sureste-noroeste y forman parte de la gran cuenca del Usumacinta (Müllerried 1957, INEGI 1988a). Asimismo, en la zona de estudio se encuentra la laguna de Nahá de origen cárstico, importante para el asentamiento poblacional de los lacandones, la cual forma parte del sistema endorreico que desagua también en la misma cuenca por medio de galerías subterráneas. El clima, según el sistema de Köppen modificado por García (1973), es cálido húmedo con lluvias abundantes en el verano y parte del otoño, y una temporada seca y corta de marzo a abril (Aw2 (w)(i’)g). La temperatura media anual es de 25 ºC y la precipitación total anual es de 2,500 mm (Muench-Navarro 1978, INEGI 1988c). Los suelos predominantes son rendzinas y litosoles, de coloración roja y gris, respectivamente; son suelos poco desarrollados que no sobrepasan 50 cm de espesor, generalmente arcillosos (Muench-Navarro 1982). Miranda (1975), quien dividió en regiones florísticas el estado de Chiapas, denominó “llanuras y declives del norte del macizo central” a la región donde se localiza la comunidad de estudio. Las condiciones climáticas y edáficas descritas favorecen el desarrollo de la selva alta y mediana perennifolia, también denominadas bosque tropical perennifolio (Miranda 1961, Miranda & Hernández-Xolocotzi 1963, Rzedowski 1978, Miranda 1975, 1976, INEGI 1988d). La comunidad de Nahá fue seleccionada en función de: 1) la existencia de áreas con vegetación madura; 2) aprovechamiento tradicional de la vegetación, diverso e intenso; 3) la aceptación de las autoridades (comunales y municipales), y de la población para realizar la investigación. Después de la aceptación del proyecto por la comunidad, se planteó la necesidad de contar con guías para realizar el trabajo de campo. Así, se conformó un equipo de trabajo con miembros de la comunidad, destacados por sus conocimientos botánicos, como informantes para la realización de los muestreos de vegetación y las recolectas botánicas.
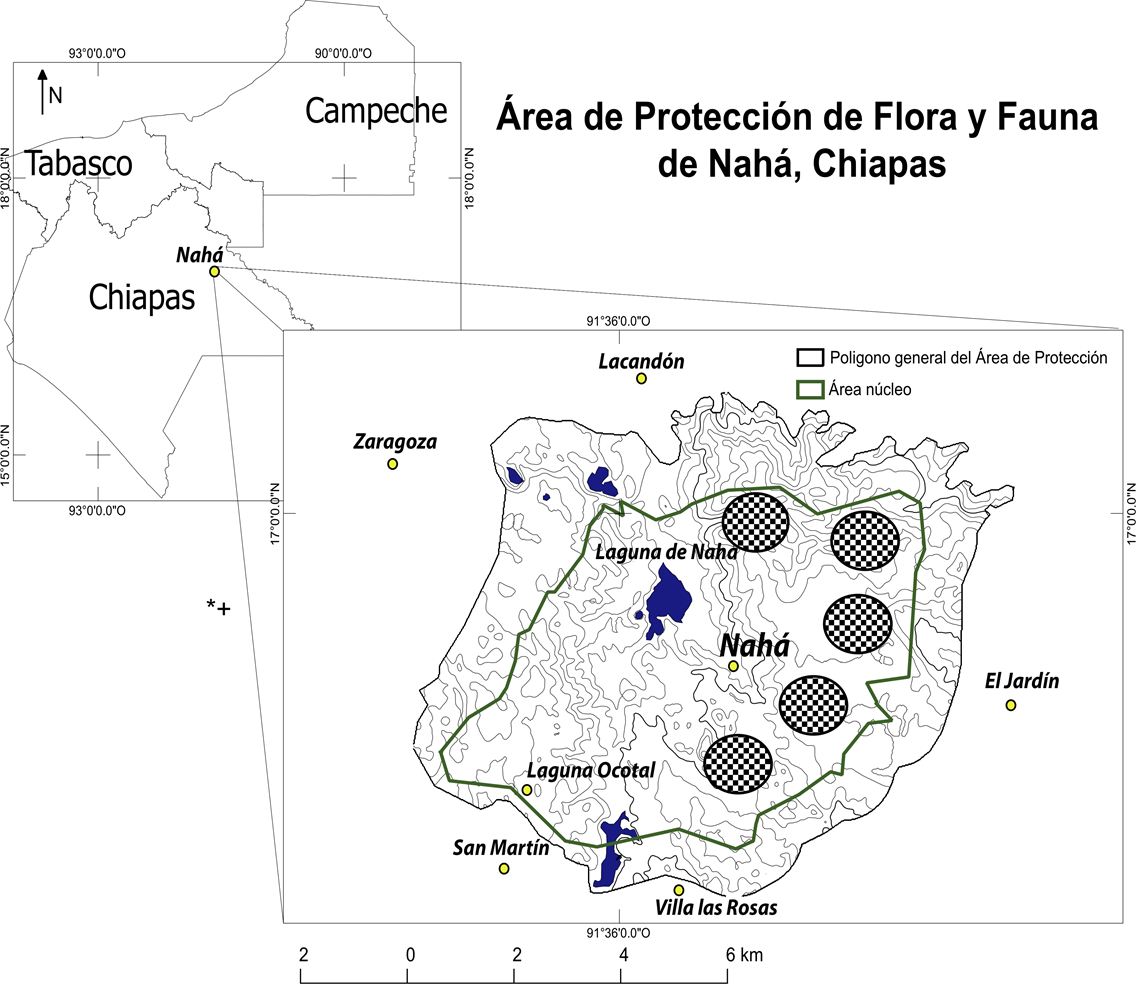
Figura 1 Localización de la comunidad de Nahá en la selva Lacandona, Chiapas, México. Los círculos indican la localización aproximada de las áreas principales de Selva Alta Perennifolia madura en que se ubicaron las 25 unidades de muestreo.
Registro de los atributos estructurales de la vegetación. Dentro de estas tierras, con ayuda de la información cartográfica de INEGI (INEGI 1988a, 1988b, 1988c, 1988d) y recorridos de campo con los informantes, se procedió a reconocer áreas con características físicas homogéneas. Así, se localizaron las áreas de bosque maduro con base en la memoria de dichos informantes quienes indicaron los sitios donde nadie recordara aprovechamiento alguno en el pasado. Para evaluar las características estructurales de la selva alta perennifolia se inventariaron 25 muestras de 400 m2 (20 × 20 m) que representa una hectárea de dichas áreas de bosque maduro (Figura 1). Los cuadros de muestreo se ubicaron sistemáticamente en un área de 2,800 ha. La distancia entre los cuadros de muestreo varió entre 0.5 y 3 km. Para facilitar el censo dentro de cada una de las parcelas, éstas se dividieron en 16 secciones. En cada muestra se realizó el registro estructural exhaustivo por sinusia (‘comunidad constituida por especies pertenecientes a un biotipo determinado de exigencias ecológicas uniformes’: Font Quer 1953) que incluye a todos los individuos maduros de cada especie, distinguiéndose de la repoblación (regeneración) natural. Con ayuda de los informantes se definió la altura máxima de cada especie, para luego proceder a la conformación de grupos de especies de acuerdo con su talla, los cuales representan los estratos reconocidos en este estudio. Los atributos de la vegetación evaluados fueron: composición florística, densidad, frecuencia, área basal (calculada con el diámetro a 1.3 m para árboles y a nivel del suelo para otras formas vitales) y altura (Müeller-Dombois & Ellenberg 1974, Bonham 1989). A cada individuo registrado se le asignó su forma vital de acuerdo con los conceptos en Font Quer (1953): árbol, arbusto, liana, trepadora, palma, herbácea perenne, hemiepífitas arbórea y arbustiva, epífita y parásita.
Los especímenes recolectados se depositaron en el herbario Isidro Palacios (SLPM) de la Universidad Autónoma de San Luis Potosí, con duplicados en el herbario MEXU, del Instituto de Biología de la Universidad Nacional Autónoma de México. La identificación se llevó a cabo principalmente en el herbario Isidro Palacios por su curador el Sr. José D. García P., con base en literatura especializada, y en algunos casos con ayuda de especialistas en algunas familias. La nomenclatura se revisó consultando la base en línea The Plant List (http://www.theplantlist.org), pero considerando en cada caso la información más aceptada por los principales especialistas de grupos. La clasificación de los helechos se basó en el sistema de Christenhusz et al. (2011) y para las angiospermas se usó el sistema APG III (APG 2009).
Resultados
Grupos taxonómicos. En el área de estudio se registraron 15,632 individuos de 283 especies de plantas vasculares que corresponden a 198 géneros, representantes de 84 familias (Apéndice 1). Los taxones registrados pertenecen a tres grupos principales. Angiospermae es el mejor representado con 273 especies, 96.47 % del total registrado, de las cuales 199 son Eudicotiledóneas, 55 Monocotiledóneas y 19 Magnólidas. Mientras que los grupos taxonómicos de Monilophyta sumaron 3.18 % de las especies totales, con nueve especies, y Gimnospermae sólo 0.35 % con una especie (Tabla 1). Las familias mejor representadas fueron Rubiaceae (9.54 %), Orchidaceae (7.42 %), Fabaceae (7.42 %), Bromeliaceae (3.89 %), Melastomataceae (3.89 %) y Lauraceae (3.18 %). Seis familias con entre nueve y 27 especies cada una, agruparon el 35.34 % de todas las especies registradas, mientras que las 78 familias restantes contribuyeron con el 64.66 % complementario.
Tabla 1 Número de taxones de la flora vascular registrados en un total de 25 muestras de 400 m2 (una hectárea), en la selva alta perennifolia de Nahá, Chiapas, México.
| Grupo taxonómico | Familias | Géneros | Especies |
|---|---|---|---|
| Monilophyta | 5 | 8 | 9 |
| Gimnospermae | 1 | 1 | 1 |
| Angiospermae | |||
| Magnólidas | 6 | 13 | 19 |
| Monocotiledóneas | 12 | 32 | 55 |
| Eudicotiledóneas | 60 | 144 | 199 |
| Total | 84 | 198 | 283 |
Estructura de la selva alta perennifolia. La selva alta perennifolia en la localidad de Nahá, Chiapas, presentó una estructura vertical compleja multiestratificada como se resume en la Tabla 2, que incluye individuos desde 0.10 como Chamaedorea pinnatifrons en el estrato herbáceo perenne a casi 60 m de altura como Swietenia macrophylla en el estrato arbóreo sobresaliente, y un área basal total de 136.78 m2. En la misma Tabla 2 se presentan los atributos estructurales y los valores de importancia (VI) para las especies por estratos. a) El estrato inferior herbáceo perenne estuvo integrado por 3,712 individuos con una altura máxima de 2.0 m, pertenecientes a 27 especies. Este estrato constituyó poco más que 20 % de la densidad total, pero sólo 0.33 % del área basal. Las especies dominantes por su densidad fueron palmas pequeñas, como Chamaedorea oblongata, C. pinnatifrons y C. elegans, y plantas propiamente herbáceas entre las que destacaron Chromolaena collina y Maranta gibba; estas cinco especies en conjunto acumularon 51 % del valor de importancia total para el estrato. Otro conjunto muy característico y frecuente de este estrato fueron las pteridófitas Adiantum tenerum, A. tetraphyllum, Didymochlaena truncatula, Macrothelypteris torresiana, Trichomanes collariatum, Christella dentata, Polypodium eperopeutes, Pecluma sp. y Campyloneurum xalapense. b) El siguiente es un estrato transicional que denominamos arbóreo-arbustivo-megafórbico, conformado por 50 especies con más que 3,200 individuos que sumaron alrededor del 2 % del área basal total. Este estrato se distingue por la presencia de arbustos y árboles de talla baja (de 2.2 a 6.5 m de altura), aunque también se encuentran algunas palmas y herbáceas perennes de gran talla o megaforbias. Entre las especies con valores de importancia más altos destacaron por su densidad las palmas Chamaedorea arenbergiana y Cryosophila stauracantha, la herbácea perenne Justicia fimbriata y el arbusto Ageratina ligustrina. Otras especies comunes fueron Heliconia librata, Ardisia nigrescens, Telanthophora grandifolia, Verbesina lanata, Exothea sp., Palicourea tetragona, Justicia borrerae y Clidemia petiolaris; que acumularon casi 50 % del valor de importancia para su estrato. c) El estrato arbóreo inferior estuvo conformado por 46 especies, casi en su totalidad de árboles pequeños y algunos arbustos altos; los individuos adultos de las especies de este estrato presentan alturas entre 7 y 14 m. Este estrato presentó la menor densidad en la selva, sólo alrededor de 1,100 individuos que representaron 7 % de la densidad total, aunque su área basal (2.9 % del total) superó a los estratos precedentes. Las especies dominantes por su densidad y área basal, con 52 % del valor de importancia del estrato, fueron Styrax warscewiczii, Miconia hyperprasina (arbusto), Garcinia intermedia, Conostegia caelestis, Siparuna thecaphora, Ocotea cernua, Polygala jamaicensis, Citharexylum hexangulare (arbusto), Rinorea hummelii y Ardisia karwinskyana. d) Entre los 15 y 23 m de altura se reconoció el estrato arbóreo medio, con 55 especies entre árboles, plantas trepadoras, lianas y algunas hemiepífitas arbóreas; en conjunto este estrato constituyó alrededor de 12 y 8 % de la densidad y del área basal global, respectivamente. Las especies más importantes fueron Trophis mexicana, Photinia microcarpa, Salacia impressifolia, Dracaena americana, Protium copal, Saurauia scabrida, Psychotria panamensis, Serjania caracasana, Miconia barbinervis, Senna racemosa y Pouteria durlandii, que suman 51 % del valor de importancia total para este estrato. En este estrato arbóreo medio se lograron registrar dos especies parásitas de la familia Santalaceae y 33 epífitas pertenecientes a las familias Orchidaceae (20 especies), Bromeliaceae (11), Piperaceae (1) y Araceae (1), mismas que por su peculiar forma vital y su ubicación únicamente se les registró su presencia e identidad taxonómica sin medirles los otros atributos estructurales (Apéndice 1). e) El estrato arbóreo alto forma un dosel continuo de entre 25 a 38 m de altura. Con 55 taxones registrados, fue el estrato con la densidad más alta, pues superó en cuatro veces la densidad registrada en el estrato arbóreo inferior y en más que dos veces la del estrato arbóreo medio. El área basal alcanzó 40 % del valor total registrado en la comunidad. Así, las sinusias de árboles, lianas, trepadoras y hemiepífitas arbóreas fueron las más comunes en este estrato y en el arbóreo medio. Las especies de mayor importancia estructural fueron Pseudolmedia glabrata, Sideroxylon salicifolium, Alchornea latifolia, Podocarpus matudae, Aspidosperma megalocarpon, Aphananthe monoica y Guarea glabra, las cuales aportaron en conjunto 51% del valor de importancia total del estrato, debido principalmente a su densidad y frecuencia. f) El estrato arbóreo sobresaliente lo conformaron únicamente 15 especies con 1,567 individuos que aportaron 10 y 46 % de la densidad y del área basal total, respectivamente. Este estrato constó de grandes árboles de entre 40 y 60 m altura y diámetro normal de 0.38 a 1.11 m, aunque algunos (1 %), particularmente de Terminalia amazonia, Swietenia macrophylla y Ficus apollinaris, llegaron a tener diámetros de 1.24 hasta 2.1 m. Así, estos individuos sobrepasan notablemente al resto de la vegetación, pero como se presentan muy distantes entre sí, su dosel es discontinuo. Las cinco especies con los valores de importancia más altos fueron T. amazonia, Dialium guianense, Billia rosea, Guatteria anomala y Dussia mexicana, y acumularon 75 % del valor de importancia total del estrato. Cabe señalar que T. amazonia presentó, por mucho, el valor de importancia absoluto y relativo más alto de toda la comunidad.
Tabla 2 Atributos estructurales y valores de importancia de las especies registradas en un total de 25 muestras de 400 m2, en la selva alta perennifolia de Nahá, Chiapas, México (*).
| Estratos/ Especies | Forma vital | Altura (m) | D A | A B A | F A | D R % | A B R % | F R % | V I | V I R % | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Máx. | Mín. | |||||||||||
| (a) Estrato herbáceo perenne | 3,712 | 4,471.50 | 23.77 | 0.32 | 11.35 | 35.44 | 100.0 | |||||
| Chamaedorea oblongata | Pa | 2.0 | 0.3 | 760 | 847.77 | 68 | 4.86 | 0.06 | 0.87 | 5.79 | 16.34 | |
| Chamaedorea pinnatifrons | Pa | 0.5 | 0.1 | 529 | 474.58 | 80 | 3.38 | 0.03 | 1.02 | 4.44 | 12.52 | |
| Chamaedorea elegans | Pa | 1.7 | 0.2 | 446 | 347.88 | 64 | 2.85 | 0.03 | 0.82 | 3.69 | 10.42 | |
| Chromolaena collina | He | 1.2 | 0.5 | 206 | 39.14 | 68 | 1.32 | 0.00 | 0.87 | 2.19 | 6.17 | |
| Maranta gibba | He | 1.0 | 0.3 | 202 | 91.48 | 68 | 1.29 | 0.01 | 0.87 | 2.17 | 6.11 | |
| Asplundia labela | Ep | 1.0 | 0.2 | 162 | 582.14 | 52 | 1.04 | 0.04 | 0.66 | 1.74 | 4.91 | |
| Clidemia setosa | He | 1.0 | 0.3 | 95 | 14.45 | 60 | 0.61 | 0.00 | 0.76 | 1.37 | 3.88 | |
| Chamaedorea metallica | Pa | 1.1 | 0.4 | 121 | 93.23 | 44 | 0.77 | 0.01 | 0.56 | 1.34 | 3.79 | |
| Adiantum tenerum | He | 2.0 | 0.5 | 126 | 483.03 | 28 | 0.81 | 0.04 | 0.36 | 1.20 | 3.38 | |
| Psychotria elata | Ab | 2.0 | 1.2 | 76 | 121.12 | 48 | 0.49 | 0.01 | 0.61 | 1.11 | 3.12 | |
| Adiantum tetraphyllum | He | 1.0 | 0.7 | 99 | 262.40 | 24 | 0.63 | 0.02 | 0.31 | 0.96 | 2.70 | |
| Spathiphyllum phryniifolium | He | 1.0 | 0.4 | 100 | 164.68 | 24 | 0.64 | 0.01 | 0.31 | 0.96 | 2.70 | |
| Olyra latifolia | He | 0.3 | 0.2 | 93 | 17.67 | 28 | 0.59 | 0.00 | 0.36 | 0.95 | 2.69 | |
| Psychotria costivenia | He | 1.0 | 0.8 | 72 | 25.77 | 32 | 0.46 | 0.00 | 0.41 | 0.87 | 2.46 | |
| Didymochlaena truncatula | He | 1.5 | 0.7 | 92 | 254.32 | 20 | 0.59 | 0.02 | 0.25 | 0.86 | 2.43 | |
| Macrothelypteris torresiana | He | 1.5 | 1.0 | 83 | 228.50 | 20 | 0.53 | 0.02 | 0.25 | 0.80 | 2.26 | |
| Trichomanes collariatum | He | 0.5 | 0.5 | 75 | 114.00 | 20 | 0.48 | 0.01 | 0.25 | 0.74 | 2.10 | |
| Neomarica gracilis | He | 0.3 | 0.3 | 68 | 12.92 | 24 | 0.44 | 0.00 | 0.31 | 0.74 | 2.09 | |
| Christella dentata | He | 1.5 | 0.5 | 59 | 188.30 | 20 | 0.38 | 0.01 | 0.25 | 0.65 | 1.82 | |
| Costus pulverulentus | He | 0.7 | 0.5 | 53 | 41.34 | 20 | 0.34 | 0.00 | 0.25 | 0.60 | 1.68 | |
| Polypodium eperopeutes | Ep | 0.4 | 0.3 | 62 | 11.66 | 12 | 0.40 | 0.00 | 0.15 | 0.55 | 1.55 | |
| Ruellia matagalpae | He | 0.5 | 0.5 | 45 | 35.10 | 20 | 0.29 | 0.00 | 0.25 | 0.55 | 1.54 | |
| Pecluma sp. | Ep | 0.3 | 0.3 | 43 | 3.01 | 16 | 0.28 | 0.00 | 0.20 | 0.48 | 1.35 | |
| Renealmia mexicana | He | 1.2 | 1.0 | 20 | 3.80 | 16 | 0.13 | 0.00 | 0.20 | 0.33 | 0.94 | |
| Campyloneurum xalapense | Ep | 0.5 | 0.3 | 18 | 12.60 | 8 | 0.12 | 0.00 | 0.10 | 0.22 | 0.62 | |
| Spigelia scabra | He | 0.3 | 0.3 | 6 | 0.42 | 4 | 0.04 | 0.00 | 0.05 | 0.09 | 0.25 | |
| Passiflora ambigua | Te | 2.0 | 2.0 | 1 | 0.19 | 4 | 0.01 | 0.00 | 0.05 | 0.06 | 0.16 | |
| (b) Estrato arbóreo-arbustivo-megafórbico | 3,238 | 32,846.81 | 20.72 | 2.40 | 22.31 | 45.4 | 100.0 | |||||
| Chamaedorea arenbergiana | Pa | 3.0 | 0.1 | 535 | 691.28 | 48 | 3.42 | 0.05 | 0.61 | 4.08 | 8.99 | |
| Cryosophila stauracantha | Pa | 6.0 | 0.5 | 348 | 7,475.34 | 92 | 2.23 | 0.55 | 1.17 | 3.94 | 8.68 | |
| Justicia fimbriata | Ar | 3.0 | 1.0 | 125 | 369.25 | 100 | 0.80 | 0.03 | 1.27 | 2.10 | 4.62 | |
| Heliconia librata | He | 2.2 | 0.4 | 131 | 1,246.34 | 64 | 0.84 | 0.09 | 0.82 | 1.74 | 3.84 | |
| Ageratina ligustrina | Ab | 5.0 | 0.4 | 111 | 593.41 | 56 | 0.71 | 0.04 | 0.71 | 1.47 | 3.23 | |
| Ardisia nigrescens | Ab | 6.0 | 1.5 | 93 | 6,134.70 | 32 | 0.59 | 0.45 | 0.41 | 1.45 | 3.19 | |
| Telanthophora grandifolia | Ar | 5.0 | 0.5 | 60 | 619.17 | 76 | 0.38 | 0.05 | 0.97 | 1.40 | 3.08 | |
| Verbesina lanata | Ar | 6.0 | 1.0 | 78 | 757.86 | 64 | 0.50 | 0.06 | 0.82 | 1.37 | 3.01 | |
| Exothea sp. | Ar | 5.0 | 1.5 | 108 | 546.37 | 48 | 0.69 | 0.04 | 0.61 | 1.34 | 2.95 | |
| Palicourea tetragona | Ar | 4.0 | 1.0 | 82 | 573.25 | 60 | 0.52 | 0.04 | 0.76 | 1.33 | 2.93 | |
| Justicia borrerae | Ab | 2.5 | 0.5 | 115 | 287.22 | 40 | 0.74 | 0.02 | 0.51 | 1.27 | 2.79 | |
| Clidemia petiolaris | He | 2.5 | 0.5 | 69 | 35.14 | 60 | 0.44 | 0.00 | 0.76 | 1.21 | 2.66 | |
| Parathesis donnell-smithii | Ar | 4.0 | 1.0 | 59 | 358.06 | 56 | 0.38 | 0.03 | 0.71 | 1.12 | 2.46 | |
| Wimmeria montana | Ar | 4.0 | 0.5 | 60 | 96.83 | 56 | 0.38 | 0.01 | 0.71 | 1.10 | 2.43 | |
| Ardisia compressa | Ar | 3.0 | 0.5 | 91 | 64.64 | 40 | 0.58 | 0.00 | 0.51 | 1.10 | 2.41 | |
| Hamelia calycosa | Ar | 4.0 | 1.8 | 79 | 358.31 | 44 | 0.51 | 0.03 | 0.56 | 1.09 | 2.40 | |
| Miconia dodecandra | Ar | 5.0 | 2.0 | 31 | 1,635.92 | 56 | 0.20 | 0.12 | 0.71 | 1.03 | 2.27 | |
| Psychotria poeppigiana | He | 3.0 | 1.0 | 45 | 194.24 | 48 | 0.29 | 0.01 | 0.61 | 0.91 | 2.01 | |
| Notopleura uliginosa | Ab | 5.0 | 0.4 | 93 | 81.73 | 24 | 0.59 | 0.01 | 0.31 | 0.91 | 2.00 | |
| Piper hispidum | Ab | 3.0 | 0.4 | 65 | 79.44 | 32 | 0.42 | 0.01 | 0.41 | 0.83 | 1.83 | |
| Croton guatemalensis | Ar | 4.0 | 1.0 | 46 | 268.06 | 40 | 0.29 | 0.02 | 0.51 | 0.82 | 1.81 | |
| Acalypha macrostachya | He | 3.0 | 1.5 | 42 | 178.68 | 40 | 0.27 | 0.01 | 0.51 | 0.79 | 1.74 | |
| Psychotria pleuropoda | Ab | 4.0 | 0.5 | 69 | 549.76 | 24 | 0.44 | 0.04 | 0.31 | 0.79 | 1.73 | |
| Astrocaryum mexicanum | Pa | 4.5 | 0.5 | 62 | 976.54 | 24 | 0.40 | 0.07 | 0.31 | 0.77 | 1.70 | |
| Hamelia longipes | Ar | 6.0 | 1.5 | 43 | 359.77 | 36 | 0.28 | 0.03 | 0.46 | 0.76 | 1.67 | |
| Columnea purpusii | Hab | 3.0 | 0.4 | 64 | 931.39 | 20 | 0.41 | 0.07 | 0.25 | 0.73 | 1.61 | |
| Miconia impetiolaris | Ab | 6.0 | 0.5 | 58 | 510.02 | 24 | 0.37 | 0.04 | 0.31 | 0.71 | 1.57 | |
| Acalypha skutchii | Ab | 5.0 | 2.0 | 39 | 1,702.09 | 24 | 0.25 | 0.12 | 0.31 | 0.68 | 1.50 | |
| Tournefortia hirsutissima | Li | 6.0 | 1.5 | 28 | 468.73 | 36 | 0.18 | 0.03 | 0.46 | 0.67 | 1.48 | |
| Psychotria pubescens | Ab | 6.0 | 1.0 | 44 | 358.64 | 28 | 0.28 | 0.03 | 0.36 | 0.66 | 1.46 | |
| Notopleura latistipula | Ab | 5.0 | 0.5 | 43 | 339.64 | 24 | 0.28 | 0.02 | 0.31 | 0.61 | 1.33 | |
| Eupatorium sp. | Ar | 6.0 | 5.0 | 14 | 196.30 | 36 | 0.09 | 0.01 | 0.46 | 0.56 | 1.24 | |
| Strychnos panamensis | Li | 6.0 | 0.5 | 42 | 108.00 | 20 | 0.27 | 0.01 | 0.25 | 0.53 | 1.17 | |
| Philodendron radiatum | Te | 5.0 | 1.0 | 22 | 17.16 | 28 | 0.14 | 0.00 | 0.36 | 0.50 | 1.10 | |
| Eugenia tikalana | Ab | 6.0 | 1.0 | 28 | 861.66 | 20 | 0.18 | 0.06 | 0.25 | 0.50 | 1.09 | |
| Begonia glabra | Te | 4.0 | 1.0 | 21 | 16.38 | 28 | 0.13 | 0.00 | 0.36 | 0.49 | 1.08 | |
| Psychotria horizontalis | Ab | 5.0 | 0.5 | 32 | 405.61 | 20 | 0.20 | 0.03 | 0.25 | 0.49 | 1.08 | |
| Drymonia strigosa | Hab | 4.0 | 1.0 | 20 | 344.67 | 20 | 0.13 | 0.03 | 0.25 | 0.41 | 0.90 | |
| Miconia fulvostellata | Ar | 4.0 | 1.5 | 30 | 777.64 | 12 | 0.19 | 0.06 | 0.15 | 0.40 | 0.88 | |
| Arachnothryx stachyoidea | Ar | 6.0 | 4.0 | 12 | 186.09 | 24 | 0.08 | 0.01 | 0.31 | 0.40 | 0.87 | |
| Arachnothryx gonzaleoides | Ar | 6.0 | 4.0 | 12 | 183.15 | 24 | 0.08 | 0.01 | 0.31 | 0.40 | 0.87 | |
| Clidemia laxiflora | Ab | 6.5 | 5.0 | 9 | 359.46 | 24 | 0.06 | 0.03 | 0.31 | 0.39 | 0.86 | |
| Sinclairia deppeana | Hab | 3.0 | 2.0 | 9 | 203.94 | 20 | 0.06 | 0.01 | 0.25 | 0.33 | 0.72 | |
| Crossopetalum parviflorum | Ar | 3.5 | 1.5 | 41 | 91.04 | 4 | 0.26 | 0.01 | 0.05 | 0.32 | 0.70 | |
| Chiococca phaenostemon | Ar | 4.0 | 3.0 | 10 | 135.81 | 16 | 0.06 | 0.01 | 0.20 | 0.28 | 0.61 | |
| Bauhinia pansamalana | Ar | 3.0 | 2.0 | 8 | 42.95 | 16 | 0.05 | 0.00 | 0.20 | 0.26 | 0.57 | |
| Thevetia ahouai | Ar | 5.0 | 2.0 | 4 | 44.16 | 12 | 0.03 | 0.00 | 0.15 | 0.18 | 0.40 | |
| Tetrapterys schiedeana | Li | 4.0 | 4.0 | 5 | 24.50 | 4 | 0.03 | 0.00 | 0.05 | 0.08 | 0.19 | |
| Mikania hookeriana | Te | 4.0 | 4.0 | 2 | 6.28 | 4 | 0.01 | 0.00 | 0.05 | 0.06 | 0.14 | |
| Gurania makoyana | Te | 3.0 | 3.0 | 1 | 0.19 | 4 | 0.01 | 0.00 | 0.05 | 0.06 | 0.13 | |
| (c) Estrato arbóreo inferior | 1,094 | 39,801.44 | 7.03 | 2.89 | 14.43 | 24.38 | 100.0 | |||||
| Styrax warscewiczii | Ar | 12.0 | 0.8 | 154 | 3,467.75 | 64 | 0.99 | 0.25 | 0.82 | 2.05 | 8.42 | |
| Miconia hyperprasina | Ab | 12.0 | 1.5 | 83 | 6,385.69 | 40 | 0.53 | 0.47 | 0.51 | 1.51 | 6.18 | |
| Garcinia intermedia | Ar | 8.0 | 4.0 | 48 | 2,608.11 | 68 | 0.31 | 0.19 | 0.87 | 1.36 | 5.59 | |
| Conostegia caelestis | Ar | 7.0 | 2.0 | 70 | 2,596.00 | 52 | 0.45 | 0.19 | 0.66 | 1.30 | 5.33 | |
| Siparuna thecaphora | Ar | 8.0 | 0.5 | 89 | 1,157.26 | 48 | 0.57 | 0.08 | 0.61 | 1.27 | 5.19 | |
| Ocotea cernua | Ar | 14.0 | 0.5 | 69 | 2,648.32 | 40 | 0.44 | 0.19 | 0.51 | 1.14 | 4.69 | |
| Polygala jamaicensis | Ar | 7.0 | 3.0 | 40 | 1,303.63 | 60 | 0.26 | 0.10 | 0.76 | 1.12 | 4.58 | |
| Citharexylum hexangulare | Ab | 9.0 | 1.5 | 69 | 1,374.93 | 40 | 0.44 | 0.10 | 0.51 | 1.05 | 4.31 | |
| Rinorea hummelii | Ar | 10.0 | 3.0 | 31 | 841.78 | 56 | 0.20 | 0.06 | 0.71 | 0.97 | 3.99 | |
| Ardisia karwinskyana | Ar | 10.0 | 1.0 | 55 | 306.13 | 44 | 0.35 | 0.02 | 0.56 | 0.93 | 3.83 | |
| Lippia myriocephala | Ab | 8.0 | 0.5 | 36 | 704.06 | 28 | 0.23 | 0.05 | 0.36 | 0.64 | 2.62 | |
| Leucaena pulverulenta | Ar | 11.0 | 4.0 | 16 | 951.91 | 36 | 0.10 | 0.07 | 0.46 | 0.63 | 2.59 | |
| Ardisia paschalis | Ar | 8.0 | 3.5 | 14 | 228.88 | 40 | 0.09 | 0.02 | 0.51 | 0.62 | 2.53 | |
| Abarema zolleriana | Ar | 7.0 | 4.0 | 23 | 1,886.44 | 24 | 0.15 | 0.14 | 0.31 | 0.59 | 2.42 | |
| Trichilia japurensis | Ar | 13.0 | 4.5 | 14 | 872.12 | 32 | 0.09 | 0.06 | 0.41 | 0.56 | 2.30 | |
| Croton xalapensis | Ar | 9.0 | 2.0 | 18 | 589.57 | 28 | 0.12 | 0.04 | 0.36 | 0.52 | 2.11 | |
| Leucaena diversifolia | Ar | 10.0 | 4.0 | 17 | 1,107.40 | 24 | 0.11 | 0.08 | 0.31 | 0.50 | 2.03 | |
| Gonzalagunia thyrsoidea | Ar | 13.0 | 4.0 | 19 | 1,204.71 | 20 | 0.12 | 0.09 | 0.25 | 0.46 | 1.90 | |
| Eugenia acapulcensis | Ab | 12.0 | 2.0 | 17 | 485.69 | 24 | 0.11 | 0.04 | 0.31 | 0.45 | 1.85 | |
| Hampea stipitata | Ar | 14.0 | 0.5 | 19 | 903.08 | 20 | 0.12 | 0.07 | 0.25 | 0.44 | 1.81 | |
| Lonchocarpus verrucosus | Ar | 10.0 | 4.0 | 20 | 802.40 | 20 | 0.13 | 0.06 | 0.25 | 0.44 | 1.81 | |
| Erythrina berteroana | Ar | 8.0 | 3.0 | 9 | 956.57 | 24 | 0.06 | 0.07 | 0.31 | 0.43 | 1.78 | |
| Calliandra tergemina var. emarginata | Ar | 14.0 | 2.5 | 14 | 441.54 | 24 | 0.09 | 0.03 | 0.31 | 0.43 | 1.75 | |
| Licaria caudata | Ar | 8.0 | 6.0 | 17 | 662.60 | 20 | 0.11 | 0.05 | 0.25 | 0.41 | 1.69 | |
| Myrcia splendens | Ar | 11.0 | 7.0 | 7 | 587.25 | 24 | 0.04 | 0.04 | 0.31 | 0.39 | 1.61 | |
| Mortoniodendron vestitum | Ar | 8.0 | 4.0 | 15 | 583.10 | 20 | 0.10 | 0.04 | 0.25 | 0.39 | 1.61 | |
| Syngonium podophyllum | Te | 10.0 | 2.0 | 15 | 280.91 | 20 | 0.10 | 0.02 | 0.25 | 0.37 | 1.52 | |
| Conostegia icosandra | Ar | 8.0 | 2.0 | 19 | 576.61 | 16 | 0.12 | 0.04 | 0.20 | 0.37 | 1.51 | |
| Psychotria flava | Ar | 8.0 | 4.0 | 7 | 219.37 | 20 | 0.04 | 0.02 | 0.25 | 0.32 | 1.29 | |
| Casearia bartlettii | Ar | 10.0 | 6.0 | 9 | 677.94 | 16 | 0.06 | 0.05 | 0.20 | 0.31 | 1.28 | |
| Croton billbergianus subsp. pyramidalis | Ar | 12.0 | 4.0 | 10 | 329.23 | 16 | 0.06 | 0.02 | 0.20 | 0.29 | 1.20 | |
| Arachnothryx capitellata | Ab | 12.0 | 10.0 | 6 | 661.86 | 16 | 0.04 | 0.05 | 0.20 | 0.29 | 1.19 | |
| Hauya elegans subsp. cornuta | Ar | 7.0 | 5.0 | 5 | 48.86 | 20 | 0.03 | 0.00 | 0.25 | 0.29 | 1.19 | |
| Ternstroemia tepezapote | Ar | 14.0 | 12.0 | 4 | 597.08 | 16 | 0.03 | 0.04 | 0.20 | 0.27 | 1.12 | |
| Hirtella americana | Ar | 7.0 | 3.0 | 4 | 38.12 | 16 | 0.03 | 0.00 | 0.20 | 0.23 | 0.95 | |
| Deherainia smaragdina | Ar | 9.0 | 7.5 | 6 | 283.50 | 12 | 0.04 | 0.02 | 0.15 | 0.21 | 0.87 | |
| Desmodium macrodesmum | Te | 13.0 | 10.0 | 6 | 2.71 | 8 | 0.04 | 0.00 | 0.10 | 0.14 | 0.58 | |
| Desmodium metallicum | Li | 12.0 | 10.0 | 3 | 80.29 | 8 | 0.02 | 0.01 | 0.10 | 0.13 | 0.52 | |
| Vanilla planifolia | Te | 8.0 | 4.0 | 5 | 3.90 | 4 | 0.03 | 0.00 | 0.05 | 0.08 | 0.34 | |
| Persea liebmannii | Ar | 10.0 | 7.0 | 3 | 81.86 | 4 | 0.02 | 0.01 | 0.05 | 0.08 | 0.31 | |
| Matayba oppositifolia | Ar | 9.0 | 4.5 | 2 | 114.86 | 4 | 0.01 | 0.01 | 0.05 | 0.07 | 0.30 | |
| Huertea cubensis | Ar | 10.0 | 10.0 | 2 | 50.26 | 4 | 0.01 | 0.00 | 0.05 | 0.07 | 0.28 | |
| Passiflora cobanensis | Te | 10.0 | 10.0 | 2 | 3.52 | 4 | 0.01 | 0.00 | 0.05 | 0.06 | 0.26 | |
| Cecropia peltata | Ar | 10.0 | 10.0 | 1 | 63.61 | 4 | 0.01 | 0.00 | 0.05 | 0.06 | 0.25 | |
| Inga belizensis | Ar | 7.0 | 7.0 | 1 | 28.27 | 4 | 0.01 | 0.00 | 0.05 | 0.06 | 0.24 | |
| Valeriana scandens | Te | 9.0 | 9.0 | 1 | 1.76 | 4 | 0.01 | 0.00 | 0.05 | 0.06 | 0.24 | |
| (d) Estrato arbóreo medio | 1,931 | 116,303.92 | 12.32 | 8.53 | 20.62 | 41.54 | 100.0 | |||||
| Trophis mexicana | Ar | 15.0 | 1.0 | 396 | 8,800.88 | 100 | 2.53 | 0.64 | 1.27 | 4.45 | 10.71 | |
| Photinia microcarpa | Ar | 20.0 | 2.0 | 180 | 14,190.29 | 64 | 1.15 | 1.04 | 0.82 | 3.00 | 7.23 | |
| Salacia impressifolia | Ar | 15.0 | 2.0 | 102 | 6,995.05 | 100 | 0.65 | 0.51 | 1.27 | 2.44 | 5.87 | |
| Dracaena americana | Ar | 18.0 | 1.5 | 89 | 6,566.22 | 84 | 0.57 | 0.48 | 1.07 | 2.12 | 5.10 | |
| Protium copal | Ar | 20.0 | 1.5 | 147 | 2,205.67 | 40 | 0.94 | 0.16 | 0.51 | 1.61 | 3.88 | |
| Saurauia scabrida | Ar | 20.0 | 1.0 | 77 | 3,374.61 | 56 | 0.49 | 0.25 | 0.71 | 1.45 | 3.50 | |
| Psychotria panamensis | Ar | 16.0 | 2.5 | 71 | 1,232.30 | 60 | 0.45 | 0.09 | 0.76 | 1.31 | 3.15 | |
| Serjania caracasana | Li | 15.0 | 8.0 | 86 | 1,617.28 | 48 | 0.55 | 0.12 | 0.61 | 1.28 | 3.08 | |
| Miconia barbinervis | Ar | 15.0 | 2.0 | 57 | 2,673.12 | 56 | 0.36 | 0.20 | 0.71 | 1.27 | 3.07 | |
| Senna racemosa | Li | 15.0 | 5.0 | 71 | 1,430.59 | 48 | 0.45 | 0.10 | 0.61 | 1.17 | 2.82 | |
| Pouteria durlandii | Li | 20.0 | 1.0 | 80 | 2,831.35 | 28 | 0.51 | 0.21 | 0.36 | 1.08 | 2.59 | |
| Cissus biformifolia | Li | 17.0 | 6.0 | 59 | 452.06 | 48 | 0.38 | 0.03 | 0.61 | 1.02 | 2.46 | |
| Clusia lundellii | Li | 15.0 | 5.0 | 34 | 372.08 | 56 | 0.22 | 0.03 | 0.71 | 0.96 | 2.31 | |
| Oecopetalum mexicanum | Ar | 23.0 | 10.0 | 27 | 4,547.75 | 32 | 0.17 | 0.33 | 0.41 | 0.91 | 2.20 | |
| Coccoloba barbadensis | Ar | 20.0 | 10.0 | 18 | 6,650.44 | 24 | 0.12 | 0.49 | 0.31 | 0.91 | 2.18 | |
| Philodendron smithii | Te | 20.0 | 3.0 | 44 | 136.25 | 48 | 0.28 | 0.01 | 0.61 | 0.90 | 2.17 | |
| Clusia guatemalensis | Har | 20.0 | 7.0 | 30 | 5,938.31 | 20 | 0.19 | 0.43 | 0.25 | 0.88 | 2.12 | |
| Capparidastrum mollicellum | Ar | 20.0 | 5.0 | 21 | 2,266.57 | 36 | 0.13 | 0.17 | 0.46 | 0.76 | 1.83 | |
| Clusia rosea | Har | 23.0 | 10.0 | 24 | 4,618.08 | 20 | 0.15 | 0.34 | 0.25 | 0.75 | 1.80 | |
| Heliocarpus appendiculatus | Ar | 20.0 | 5.0 | 8 | 4,525.70 | 28 | 0.05 | 0.33 | 0.36 | 0.74 | 1.78 | |
| Trichilia quadrijuga subsp. cinerascens | Ar | 18.0 | 5.0 | 22 | 2,209.62 | 32 | 0.14 | 0.16 | 0.41 | 0.71 | 1.71 | |
| Cymbopetalum penduliflorum | Ar | 22.0 | 5.0 | 13 | 1,945.95 | 36 | 0.08 | 0.14 | 0.46 | 0.68 | 1.65 | |
| Hirtella triandra | Ar | 22.0 | 10.0 | 11 | 1,901.41 | 32 | 0.07 | 0.14 | 0.41 | 0.62 | 1.49 | |
| Heliocarpus donnellsmithii | Ar | 20.0 | 7.0 | 8 | 2,284.50 | 24 | 0.05 | 0.17 | 0.31 | 0.52 | 1.26 | |
| Nectandra salicifolia | Ar | 20.0 | 15.0 | 4 | 3,915.99 | 16 | 0.03 | 0.29 | 0.20 | 0.52 | 1.24 | |
| Inga sp. | Ar | 20.0 | 12.0 | 6 | 2,585.70 | 20 | 0.04 | 0.19 | 0.25 | 0.48 | 1.16 | |
| Ilex costaricensis | Ar | 23.0 | 6.5 | 7 | 1,093.56 | 28 | 0.04 | 0.08 | 0.36 | 0.48 | 1.16 | |
| Bauhinia rubeleruziana | Ar | 18.0 | 1.5 | 10 | 1,519.11 | 24 | 0.06 | 0.11 | 0.31 | 0.48 | 1.16 | |
| Zanthoxylum juniperinum | Ar | 20.0 | 6.0 | 8 | 1,641.06 | 24 | 0.05 | 0.12 | 0.31 | 0.48 | 1.15 | |
| Licaria peckii | Li | 20.0 | 5.0 | 19 | 628.02 | 24 | 0.12 | 0.05 | 0.31 | 0.47 | 1.14 | |
| Licaria excelsa | Ar | 15.0 | 12.0 | 20 | 3,280.40 | 8 | 0.13 | 0.24 | 0.10 | 0.47 | 1.13 | |
| Cissus alata | Li | 20.0 | 6.0 | 30 | 124.12 | 20 | 0.19 | 0.01 | 0.25 | 0.46 | 1.10 | |
| Lonchocarpus rugosus | Ar | 21.0 | 6.0 | 12 | 597.74 | 24 | 0.08 | 0.04 | 0.31 | 0.43 | 1.03 | |
| Oreopanax guatemalensis | Har | 18.0 | 4.0 | 12 | 732.91 | 20 | 0.08 | 0.05 | 0.25 | 0.39 | 0.93 | |
| Garcia nutans | Ar | 15.0 | 8.0 | 10 | 1,555.85 | 16 | 0.06 | 0.11 | 0.20 | 0.38 | 0.92 | |
| Ocotea sinuata | Ar | 20.0 | 10.0 | 6 | 483.84 | 24 | 0.04 | 0.04 | 0.31 | 0.38 | 0.91 | |
| Rhynchosia erythrinoides | Li | 20.0 | 13.0 | 16 | 180.38 | 20 | 0.10 | 0.01 | 0.25 | 0.37 | 0.89 | |
| Stenanona stenopetala | Ar | 21.0 | 8.0 | 7 | 773.20 | 20 | 0.04 | 0.06 | 0.25 | 0.36 | 0.86 | |
| Marcgravia mexicana | Li | 20.0 | 10.0 | 12 | 138.97 | 20 | 0.08 | 0.01 | 0.25 | 0.34 | 0.82 | |
| Clusia salvinii | Har | 20.0 | 0.8 | 12 | 119.50 | 20 | 0.08 | 0.01 | 0.25 | 0.34 | 0.82 | |
| Trema micrantha | Ar | 23.0 | 12.0 | 5 | 1,204.97 | 16 | 0.03 | 0.09 | 0.20 | 0.32 | 0.78 | |
| Psychotria galeottiana | Ar | 15.0 | 4.5 | 7 | 280.94 | 20 | 0.04 | 0.02 | 0.25 | 0.32 | 0.77 | |
| Pimenta dioica | Ar | 20.0 | 6.5 | 5 | 779.60 | 16 | 0.03 | 0.06 | 0.20 | 0.29 | 0.70 | |
| Casearia aculeata | Ar | 19.0 | 11.0 | 5 | 582.16 | 16 | 0.03 | 0.04 | 0.20 | 0.28 | 0.67 | |
| Inga punctata | Ar | 18.0 | 1.0 | 11 | 583.83 | 12 | 0.07 | 0.04 | 0.15 | 0.27 | 0.64 | |
| Nectandra coriacea | Ar | 20.0 | 15.0 | 4 | 431.55 | 16 | 0.03 | 0.03 | 0.20 | 0.26 | 0.63 | |
| Clethra suaveolens | Ar | 15.0 | 10.0 | 4 | 631.45 | 12 | 0.03 | 0.05 | 0.15 | 0.22 | 0.54 | |
| Coccoloba cozumelensis | Ar | 20.0 | 14.0 | 4 | 502.04 | 12 | 0.03 | 0.04 | 0.15 | 0.22 | 0.52 | |
| Faramea occidentalis | Ar | 16.0 | 14.0 | 3 | 486.15 | 12 | 0.02 | 0.04 | 0.15 | 0.21 | 0.50 | |
| Philodendron sagittifolium | Te | 15.0 | 10.0 | 8 | 6.24 | 12 | 0.05 | 0.00 | 0.15 | 0.20 | 0.49 | |
| Strychnos brachistantha | Li | 20.0 | 7.0 | 3 | 22.96 | 12 | 0.02 | 0.00 | 0.15 | 0.17 | 0.42 | |
| Tonduzia longifolia | Ar | 19.0 | 15.0 | 2 | 906.34 | 4 | 0.01 | 0.07 | 0.05 | 0.13 | 0.31 | |
| Cecropia obtusifolia | Ar | 18.0 | 15.0 | 2 | 208.12 | 8 | 0.01 | 0.02 | 0.10 | 0.13 | 0.31 | |
| Amphitecna silvicola | Ar | 18.0 | 18.0 | 1 | 314.16 | 4 | 0.01 | 0.02 | 0.05 | 0.08 | 0.19 | |
| Rogiera stenosiphon | Ar | 16.0 | 16.0 | 1 | 226.98 | 4 | 0.01 | 0.02 | 0.05 | 0.07 | 0.18 | |
| (e) Estrato arbóreo alto | 4,090 | 539,404.02 | 26.23 | 39.40 | 22.46 | 88.09 | 100.0 | |||||
| Pseudolmedia glabrata | Ar | 35.0 | 0.5 | 570 | 72,000.43 | 100 | 3.65 | 5.26 | 1.27 | 10.18 | 11.56 | |
| Sideroxylon salicifolium | Ar | 25.0 | 0.5 | 1109 | 24,687.95 | 80 | 7.09 | 1.80 | 1.02 | 9.92 | 11.26 | |
| Alchornea latifolia | Ar | 35.0 | 2.0 | 647 | 24,746.76 | 80 | 4.14 | 1.81 | 1.02 | 6.97 | 7.91 | |
| Podocarpus matudae | Ar | 25.0 | 0.5 | 225 | 61,477.44 | 40 | 1.44 | 4.49 | 0.51 | 6.44 | 7.32 | |
| Aspidosperma megalocarpon | Ar | 37.0 | 30.0 | 16 | 49,151.83 | 44 | 0.10 | 3.59 | 0.56 | 4.26 | 4.83 | |
| Aphananthe monoica | Ar | 30.0 | 6.0 | 71 | 30,289.61 | 68 | 0.45 | 2.21 | 0.87 | 3.54 | 4.01 | |
| Guarea glabra | Ar | 32.0 | 0.5 | 216 | 10,775.38 | 60 | 1.38 | 0.79 | 0.76 | 2.93 | 3.33 | |
| Quercus lancifolia | Ar | 37.0 | 3.0 | 54 | 22,880.28 | 40 | 0.35 | 1.67 | 0.51 | 2.53 | 2.87 | |
| Calophyllum brasiliense | Ar | 30.0 | 0.5 | 87 | 12,918.16 | 64 | 0.56 | 0.94 | 0.82 | 2.32 | 2.63 | |
| Cedrela odorata | Ar | 30.0 | 27.0 | 8 | 19,574.34 | 32 | 0.08 | 1.43 | 0.40 | 1.91 | 2.17 | |
| Schultesianthus leucanthus | Li | 30.0 | 15.0 | 41 | 8,854.35 | 64 | 0.26 | 0.65 | 0.82 | 1.73 | 1.96 | |
| Manilkara zapota | Ar | 36.0 | 0.5 | 125 | 6,529.85 | 28 | 0.80 | 0.48 | 0.36 | 1.63 | 1.85 | |
| Pouteria campechiana | Ar | 30.0 | 0.5 | 156 | 1,854.16 | 32 | 1.00 | 0.14 | 0.41 | 1.54 | 1.75 | |
| Wimmeria bartlettii | Ar | 25.0 | 0.4 | 134 | 1,968.99 | 32 | 0.86 | 0.14 | 0.41 | 1.41 | 1.60 | |
| Fraxinus uhdei | Ar | 34.0 | 6.0 | 37 | 9,204.72 | 36 | 0.24 | 0.67 | 0.46 | 1.37 | 1.55 | |
| Mosquitoxylum jamaicense | Ar | 30.0 | 15.0 | 10 | 10,660.80 | 40 | 0.06 | 0.78 | 0.51 | 1.35 | 1.54 | |
| Ulmus mexicana | Ar | 38.0 | 6.0 | 11 | 10,416.36 | 40 | 0.07 | 0.76 | 0.51 | 1.34 | 1.52 | |
| Hillia tetrandra | Har | 25.0 | 1.2 | 34 | 10,651.50 | 24 | 0.22 | 0.78 | 0.31 | 1.30 | 1.48 | |
| Coccoloba hondurensis | Ar | 30.0 | 18.0 | 6 | 14,292.76 | 16 | 0.04 | 1.04 | 0.20 | 1.29 | 1.46 | |
| Pleuranthodendron lindenii | Ar | 28.0 | 2.0 | 30 | 8,994.28 | 32 | 0.19 | 0.66 | 0.41 | 1.26 | 1.43 | |
| Tabebuia rosea | Ar | 27.0 | 23.0 | 4 | 14,397.24 | 12 | 0.03 | 1.05 | 0.15 | 1.23 | 1.40 | |
| Genipa americana | Ar | 25.0 | 11.0 | 14 | 6,974.25 | 40 | 0.09 | 0.51 | 0.51 | 1.11 | 1.26 | |
| Pseudobombax ellipticum | Ar | 30.0 | 8.0 | 7 | 9,219.70 | 28 | 0.04 | 0.67 | 0.36 | 1.08 | 1.22 | |
| Topobea laevigata | Har | 30.0 | 10.0 | 28 | 8,636.13 | 20 | 0.18 | 0.63 | 0.25 | 1.07 | 1.21 | |
| Morinda panamensis | Ar | 27.0 | 6.0 | 9 | 8,182.67 | 32 | 0.06 | 0.60 | 0.41 | 1.06 | 1.21 | |
| Mabea excelsa | Ar | 30.0 | 6.5 | 21 | 7,299.36 | 28 | 0.13 | 0.53 | 0.36 | 1.02 | 1.16 | |
| Chrysophyllum mexicanum | Ar | 25.0 | 22.0 | 3 | 11,498.20 | 12 | 0.02 | 0.84 | 0.15 | 1.01 | 1.15 | |
| Monstera deliciosa | Te | 30.0 | 5.0 | 46 | 245.56 | 48 | 0.29 | 0.02 | 0.61 | 0.92 | 1.05 | |
| Chionanthus ligustrinus | Ar | 27.0 | 6.0 | 15 | 6,177.84 | 28 | 0.10 | 0.45 | 0.36 | 0.90 | 1.03 | |
| Fridericia schumanniana | Li | 35.0 | 15.0 | 33 | 2,897.96 | 36 | 0.21 | 0.21 | 0.46 | 0.88 | 1.00 | |
| Dendropanax arboreus | Ar | 25.0 | 9.0 | 12 | 4,761.24 | 32 | 0.08 | 0.35 | 0.41 | 0.83 | 0.95 | |
| Inga pavoniana | Ar | 25.0 | 25.0 | 1 | 10,025.00 | 4 | 0.01 | 0.73 | 0.05 | 0.79 | 0.90 | |
| Nectandra membranacea | Ar | 35.0 | 21.0 | 7 | 7,606.73 | 12 | 0.04 | 0.56 | 0.15 | 0.75 | 0.86 | |
| Disciphania calocarpa | Li | 30.0 | 23.0 | 38 | 431.96 | 36 | 0.24 | 0.03 | 0.46 | 0.73 | 0.83 | |
| Gouania lupuloides | Li | 30.0 | 18.0 | 23 | 537.26 | 40 | 0.15 | 0.04 | 0.51 | 0.70 | 0.79 | |
| Vitis bourgaeana | Te | 35.0 | 18.0 | 25 | 980.41 | 36 | 0.16 | 0.07 | 0.46 | 0.69 | 0.78 | |
| Magnolia schiedeana | Ar | 25.0 | 12.0 | 8 | 3,144.13 | 32 | 0.05 | 0.23 | 0.41 | 0.69 | 0.78 | |
| Cojoba arborea | Ar | 25.0 | 6.0 | 15 | 4,073.57 | 20 | 0.10 | 0.30 | 0.25 | 0.65 | 0.74 | |
| Philodendron standleyi | Te | 30.0 | 10.0 | 23 | 169.11 | 36 | 0.15 | 0.01 | 0.46 | 0.62 | 0.70 | |
| Celastrus vulcanicolus | Li | 35.0 | 20.0 | 23 | 647.06 | 32 | 0.15 | 0.05 | 0.41 | 0.60 | 0.68 | |
| Semialarium mexicanum | Li | 25.0 | 10.0 | 30 | 581.26 | 28 | 0.19 | 0.04 | 0.36 | 0.59 | 0.67 | |
| Guettarda combsii | Ar | 30.0 | 9.0 | 4 | 4,419.60 | 16 | 0.03 | 0.32 | 0.20 | 0.55 | 0.63 | |
| Compsoneura sprucei | Ar | 25.0 | 5.0 | 12 | 1,424.66 | 28 | 0.08 | 0.10 | 0.36 | 0.54 | 0.61 | |
| Bursera simaruba | Ar | 27.0 | 6.0 | 11 | 2,135.67 | 24 | 0.07 | 0.16 | 0.31 | 0.53 | 0.60 | |
| Trichospermum mexicanum | Ar | 31.0 | 20.0 | 3 | 3,955.67 | 12 | 0.02 | 0.29 | 0.15 | 0.46 | 0.52 | |
| Securidaca diversifolia | Li | 30.0 | 12.0 | 22 | 391.22 | 20 | 0.14 | 0.03 | 0.25 | 0.42 | 0.48 | |
| Rhynchosia pyramidalis | Li | 30.0 | 25.0 | 18 | 364.34 | 20 | 0.12 | 0.03 | 0.25 | 0.40 | 0.45 | |
| Heteropterys cotinifolia | Li | 30.0 | 10.0 | 12 | 877.22 | 20 | 0.08 | 0.06 | 0.25 | 0.40 | 0.45 | |
| Orthion malpighiifolium | Ar | 25.0 | 18.0 | 4 | 1,626.55 | 16 | 0.03 | 0.12 | 0.20 | 0.35 | 0.40 | |
| Passiflora cookii | Te | 25.0 | 15.0 | 15 | 15.62 | 16 | 0.10 | 0.00 | 0.20 | 0.30 | 0.34 | |
| Margaritaria nobilis | Ar | 25.0 | 25.0 | 2 | 2,166.06 | 8 | 0.01 | 0.16 | 0.10 | 0.27 | 0.31 | |
| Mascagnia dipholiphylla | Li | 25.0 | 18.0 | 6 | 93.81 | 16 | 0.04 | 0.01 | 0.20 | 0.25 | 0.28 | |
| Palicourea domingensis | Ar | 30.0 | 20.0 | 3 | 1,480.91 | 8 | 0.02 | 0.11 | 0.10 | 0.23 | 0.26 | |
| Serjania atrolineata | Li | 30.0 | 16.0 | 5 | 23.54 | 12 | 0.03 | 0.00 | 0.15 | 0.19 | 0.21 | |
| Doliocarpus dentatus | Li | 25.0 | 25.0 | 1 | 12.56 | 4 | 0.01 | 0.00 | 0.05 | 0.06 | 0.07 | |
| (f) Estrato arbóreo sobresaliente | 1,567 | 634,971.40 | 10.02 | 46.43 | 8.67 | 65.10 | 100.0 | |||||
| Terminalia amazonia | Ar | 57.0 | 11.0 | 39 | 373,579.50 | 100 | 0.25 | 27.31 | 1.27 | 28.84 | 44.29 | |
| Dialium guianense | Ar | 42.0 | 1.5 | 662 | 25,581.16 | 84 | 4.23 | 1.87 | 1.07 | 7.18 | 11.02 | |
| Billia rosea | Ar | 50.0 | 0.5 | 208 | 47,534.48 | 48 | 1.33 | 3.48 | 0.61 | 5.42 | 8.32 | |
| Guatteria anomala | Ar | 49.0 | 2.0 | 91 | 36,152.03 | 40 | 0.58 | 2.64 | 0.51 | 3.73 | 5.74 | |
| Dussia mexicana | Ar | 40.0 | 1.0 | 356 | 10,917.94 | 48 | 2.28 | 0.80 | 0.61 | 3.69 | 5.66 | |
| Virola guatemalensis | Ar | 42.0 | 3.0 | 87 | 29,951.07 | 64 | 0.56 | 2.19 | 0.82 | 3.56 | 5.47 | |
| Quercus skinneri | Ar | 40.0 | 10.0 | 35 | 23,087.61 | 36 | 0.22 | 1.69 | 0.46 | 2.37 | 3.64 | |
| Swietenia macrophylla | Ar | 60.0 | 4.0 | 6 | 27,492.23 | 24 | 0.04 | 2.01 | 0.31 | 2.35 | 3.62 | |
| Ficus apollinaris | Har | 40.0 | 11.0 | 22 | 15,180.03 | 44 | 0.14 | 1.11 | 0.56 | 1.81 | 2.78 | |
| Vochysia guatemalensis | Ar | 40.0 | 6.0 | 14 | 12,608.42 | 52 | 0.09 | 0.92 | 0.66 | 1.67 | 2.57 | |
| Ficus pertusa | Har | 40.0 | 9.0 | 12 | 6,492.65 | 36 | 0.08 | 0.47 | 0.46 | 1.01 | 1.55 | |
| Magnolia grandiflora | Ar | 40.0 | 15.0 | 8 | 8,854.06 | 24 | 0.05 | 0.65 | 0.31 | 1.00 | 1.54 | |
| Chione venosa | Ar | 50.0 | 11.0 | 8 | 5,467.05 | 32 | 0.05 | 0.40 | 0.41 | 0.86 | 1.32 | |
| Bernoullia flammea | Ar | 45.0 | 6.0 | 7 | 7,322.07 | 20 | 0.04 | 0.54 | 0.25 | 0.83 | 1.28 | |
| Coussapoa oligocephala | Har | 40.0 | 25.0 | 12 | 4,751.10 | 28 | 0.08 | 0.35 | 0.36 | 0.78 | 1.20 | |
| Total | 15,632 | 1,367,799.09 | 100.0 | 100.0 | 100.0 | 300.00 | ||||||
* Las formas vitales fueron: árbol (Ar); arbusto (Ab); epífita (Ep); hemiepífita arbórea (Har); hemiepífita arbustiva (Hab); herbácea perenne (He); liana (Li); palma (Pa); trepadora (Te).
DA: densidad absoluta (individuos/ha); ABA: área basal absoluta (cm2/ha);
FA: frecuencia absoluta; DR: densidad relativa (%); ABR: área basal relativa (%);
FR: frecuencia relativa (%); VI: valor de importancia, suma de los valores de DR, ABR, y FR;
VIR: valor de importancia relativa, correspondiente al estrato (%).
Atributos estructurales por estrato y formas vitales. En relación con los atributos estructurales por estrato, la densidad y el valor de importancia variaron entre estratos en forma inconsistente e independiente; en cambio el área basal creció en forma constante desde el estrato herbáceo al sobresaliente (Tabla 3). La densidad fue marcadamente mayor en los estratos herbáceo y arbóreo-arbustivo-megafórbico que en los estratos arbóreos, con la excepción notable del estrato arbóreo alto, donde se registraron poco más que 4,000 ind/ha, el mayor número por estrato con especies como Sideroxylon salicifolium, Alchornea latifolia y Pseudolmedia glabrata con densidades entre 500 y 1,000 ind/ha. Por su parte, los estratos herbáceo y el arbóreo-arbustivo-megafórbico tuvieron altas densidades debido en gran medida a la abundancia de palmas; así, en el estrato herbáceo, sólo las especies de Chamaedorea aportaron 1,735 ind/ha, y en el estrato arbóreo-arbustivo-megafórbico para Chamaedorea arenbergiana y Cryosophila stauracantha se registraron 535 y 348 ind/ha, respectivamente. En el estrato sobresaliente sus especies presentaron densidades bajas (con excepción de Dialium guianense, Dussia mexicana y Billia rosea, con fuerte repoblación), pero individuos de gran corpulencia, tanto en altura como en área basal; así, sólo Terminalia amazonia con 39 individuos aportó 27 % del área basal total en la comunidad.
Tabla 3 Riqueza de especies, densidad, área basal y valor de importancia por estratos estimados a partir de 25 muestras de 400 m2, en la selva alta perennifolia de Náha, Chiapas, México.
| Estrato | Especies | Densidad (ind/ha) | Área basal (cm2/ha) | Valor de importancia | ||||
|---|---|---|---|---|---|---|---|---|
| Absoluto | % | Absoluta | % | Absoluta | % | Absoluto | % | |
| Herbáceo perenne | 27 | 10.88 | 3,712 | 23.75 | 4,471.50 | 0.33 | 35.44 | 11.81 |
| Arbóreo-arbustivo-megafórbico | 50 | 20.16 | 3,238 | 20.71 | 32,846.81 | 2.40 | 45.44 | 15.15 |
| Arbóreo inferior | 46 | 18.55 | 1,094 | 7.00 | 39,801.44 | 2.91 | 24.38 | 8.13 |
| Arbóreo medio | 55 | 22.18 | 1,931 | 12.35 | 116,303.92 | 8.50 | 41.55 | 13.85 |
| Arbóreo alto | 55 | 22.18 | 4,090 | 26.16 | 539,404.02 | 39.44 | 88.08 | 29.36 |
| Arbóreo sobresaliente | 15 | 6.05 | 1,567 | 10.03 | 634,971.40 | 46.42 | 65.11 | 21.70 |
| Total | 248 | 100.0 | 15,632 | 100.00 | 1,367,799.09 | 100.00 | 300.00 | 100.00 |
En la Tabla 4 se presentan los valores absolutos de densidad, área basal y el valor de importancia para el total de especies agrupadas en cada forma vital, predominando los árboles sobre las demás sinusias en todos los atributos. Así, en los árboles se registró la mayor riqueza de especies y ellos aportaron un poco más que 50 y 90 % de la densidad y área basal global, respectivamente. Otras formas vitales peculiares que caracterizan la estructura y fisonomía de las selvas fueron relativamente importantes como son las palmas (estípites), que con sólo siete especies y menos de 1 % del área basal total, fueron la segunda sinusia con mayor densidad. Las herbáceas perennes, los arbustos y las lianas les siguen en densidad y por consiguiente en valor de importancia. Las hemiepífitas arbóreas presentaron el segundo valor más alto de área basal, aunque con relativamente muy pocos individuos. Las formas vitales con menor valor de importancia fueron las trepadoras, las epífitas y las hemiepífitas arbustivas.
Tabla 4 Riqueza de especies, densidad, área basal y valor de importancia por formas vitales estimadas a partir de 25 muestras de 400 m2, en selva alta perennifolia de Náha, Chiapas, México.
| Formas vitales | Especies | Densidad (ind/ha) | Área basal (cm2/ha) | Valor de importancia | ||||
|---|---|---|---|---|---|---|---|---|
| Absoluto | % | Absoluta | % | Absoluta | % | Absoluta | % | |
| Árbol | 143 | 57.66 | 8,423 | 53.88 | 1,245,976.34 | 91.09 | 205.47 | 68.49 |
| Palma | 7 | 2.82 | 2,801 | 17.92 | 10,906.62 | 0.80 | 24.07 | 8.02 |
| Herbácea perenne | 21 | 8.47 | 1,781 | 11.39 | 3,631.72 | 0.27 | 20.68 | 6.89 |
| Arbusto | 19 | 7.66 | 1,086 | 6.95 | 21,996.73 | 1.61 | 15.79 | 5.26 |
| Liana | 26 | 10.48 | 740 | 4.73 | 24,191.87 | 1.77 | 15.68 | 5.23 |
| Hemiepífita arbórea | 9 | 3.63 | 186 | 1.19 | 57,120.21 | 4.18 | 8.32 | 2.77 |
| Trepadora | 16 | 6.45 | 237 | 1.52 | 1,886.19 | 0.14 | 5.53 | 1.84 |
| Epífita | 4 | 1.61 | 285 | 1.82 | 609.41 | 0.04 | 2.99 | 1.00 |
| Hemiepífita arbustiva | 3 | 1.21 | 93 | 0.59 | 1,480.00 | 0.11 | 1.47 | 0.49 |
| Total | 248 | 100.00 | 15,632 | 100.00 | 1,367,799.09 | 100.00 | 300.00 | 100.00 |
Discusión
Las familias botánicas registradas en el área de estudio corresponden a las familias mejor representadas en las selvas altas perennifolias estudiadas en México por Meave del Castillo (1983) y Bongers et al. (1988), así como en las selvas neotropicales descritas por Gentry (1990), aunque su importancia relativa es algo diferente en cada localidad analizada. El alto porcentaje de familias con una o pocas especies, al parecer es una tendencia general que ha sido señalada en trabajos previos desarrollados en selvas de México y de otras partes del mundo (Sarukhán-Kermes 1968, Meave del Castillo 1983, Hubbell & Foster 1983, Gentry 1990), y parece propia de ecosistemas maduros y complejos (Margalef 1980, Odum 1985, Margalef 1998).
La complejidad en la estructura vertical de las selvas altas perennifolias da la impresión de un gradiente de estatura casi continuo desde el sotobosque hasta el dosel (donde, aunque predominan árboles perennifolios existen también algunos brevemente caducifolios). Sarukhán-Kermes. (1968), en las selvas dominadas por Terminalia amazonia de la planicie costera del Golfo de México y Meave del Castillo (1983) en la selva Lacandona, Chiapas, reconocieron tres y cuatro estratos arbóreos, respectivamente, en función de la altura media y máxima de los individuos de sus especies; ambos autores señalan que dichos estratos pudieron ser artificiales, pero los consideraron necesarios para reconocer en función del índice de dominancia, los cambios en la estratificación vertical. Posiblemente esta complejidad de la selva alta condujo a Popma et al. (1988) a distinguir sólo el sotobosque y una selva no estratificada en su estudio en la región de los Tuxtlas, Veracruz. En el presente trabajo, se pudo reconocer en el campo, gracias al profundo conocimiento de los lacandones, la existencia de seis estratos bien definidos: uno herbáceo, otro arbustivo y cuatro arbóreos con composición florística y formas vitales predominantes distintivas para cada uno.
Los atributos estructurales evaluados en este estudio permitieron estimar la dominancia de las especies en la comunidad y su contribución relativa en los estratos definidos. Los atributos estructurales implicados en el cálculo del índice de valor de importancia tuvieron diferente peso en la determinación de las especies dominantes según los estratos. Así, en el estrato herbáceo, los valores de densidad determinan la jerarquización de las especies de acuerdo con su valor de importancia; en dicho estrato, entonces, las especies con altas densidades son las que ejercen la dominancia. Para las especies de los estratos superiores, con excepción de aquellas con tasas altas de repoblación, su valor de importancia estuvo determinado por el área basal (estimador de biomasa), aunque presentan densidades bajas. En los estratos intermedios el principal atributo que les confirió a las especies la dominancia o valores altos de importancia no sólo fue el área basal, pues hubo especies con densidades no muy altas, sino de frecuencia alta (distribución espacial equidistante), como fueron los casos de Justicia fimbriata, Garcinia intermedia, Salacia impressifolia y Dracaena americana, y en otros casos, el atributo que determinó la dominancia en sus estratos respectivos sí fue la densidad de sus poblaciones (Chamaedorea arenbergiana, Cryosophila stauracantha, Styrax warscewiczii y Trophis mexicana).
A pesar de la tendencia de una densidad marcadamente mayor en los estratos herbáceo y arbóreo-arbustivo-megafórbico, el estrato arbóreo alto registró la mayor densidad por estrato. La presencia de especies como Sideroxylon salicifolium, Alchornea latifolia y Pseudolmedia glabrata con densidades entre 500 y 1,000 ind/ha explica de forma contundente el incremento de la densidad en dicho estrato. Además, S. salicifolium fue la especie con la mayor densidad poblacional en toda la comunidad. No obstante su mayor densidad, el estrato herbáceo presentó una cubierta más dispersa que la del estrato arbóreo-arbustivo-megafórbico, lo cual se apreció como tendencia creciente en los estratos subsiguientes, a menor densidad, mayor tamaño. En efecto, el área basal fue mayor en los estratos arbóreos, particularmente en el sobresaliente que representó 46 % del área basal global, y con una tendencia bien marcada a disminuir hasta ser menos que 1 % del total en el estrato herbáceo. Esta relación inversa encontrada entre la densidad y área basal es concordante con la consignada para otras selvas del país (Sarukhán Kermes 1968, Meave del Castillo 1983, Popma et al. 1988). Así, pareciera que el área basal, al ser un estimador de la biomasa, resultó mejor indicador general de dominancia que el propio valor de importancia absoluto.
El censo completo de los individuos presentes en el total de muestras estudiadas (1 ha), permitió reconocer una amplia gama de formas vitales características de las comunidades vegetales de las regiones cálido-húmedas, con 15,632 individuos que van desde 0.10 m de altura (Chamaedorea pinnatifrons) en el estrato herbáceo perenne a 60 m (Swietenia macrophylla) en el estrato arbóreo sobresaliente, y un área basal total de 136.78 m2. El número de individuos y el valor de área total basal registrado en este estudio son altos si se comparan con los presentados por Meave del Castillo (1983) y por Bongers et al. (1988) para México, o por Rajkumar & Parthasarathy (2008) para las islas Andaman, pero se trata de estimaciones que varían fuertemente en dependencia de los métodos empleados en cada estudio, particularmente del tamaño mínimo de cada individuo para incluirse en el censo y del tamaño de la muestra empleada, aunque las condiciones ambientales sí sean comparables. Por ello, Bongers et al. (1988) enfatizan que la única manera de comparar selvas de una forma objetiva es utilizar los mismos métodos.
Por otro lado, las especies consignadas como dominantes de cada estrato de la selva en Nahá han sido ampliamente reconocidas por varios autores como propias o características de rodales de selva madura (Miranda 1961, Miranda & Hernández-Xolocotzi 1963, Sarukhán-Kermes 1968, Breedlove 1973, Miranda 1975, 1976, Rzedowki 1978, 1981, Meave del Castillo 1983, Levy-Tacher 2000). Así, Miranda (1961) en el valle del Censo-Monte Líbano y Levy-Tacher (2000) en Lacanhá Chansayab, describen detalladamente algunas variantes o asociaciones de la selva alta perennifolia en la región Lacandona y ratifican la tendencia de presencia de especies dominantes o propias de vegetación madura o bien conservada como Senna racemosa, Trophis mexicana, Rinorea hummelii, Siparuna thecaphora, Dracaena americana, Hampea stipitata, Cymbopetalum penduliflorum, Salacia impressifolia, Astrocaryum mexicanum, Ardisia paschalis, Pseudolmedia glabrata, Calophyllum brasiliense, Alchornea latifolia, Inga pavoniana, Guarea glabra, Aspidosperma megalocarpon, Sideroxylon salicifolium, Swietenia macrophylla, Guatteria anomala, Dialium guianense y Terminalia amazonia.
Previamente, se han registrado formas vitales peculiares y relativamente importantes que caracterizan la estructura y fisonomía de las selvas (Richards 1952, Braun-Blanquet 1979, Ehrendorfer 1986, Bongers et al. 1988). El caso de las palmas como la segunda sinusia con mayor densidad de la selva en Nahá se explica principalmente por el alto número de individuos de las especies de Chamaedorea. Asimismo, las hemiepífitas arbóreas como la segunda sinusia con mayor valor de área basal se debe a su forma peculiar de crecimiento y morfología del tronco. En la selva de Nahá las plantas parásitas están poco representadas; en cambio, las epífitas son muy abundantes y se distribuyen a través de la mayoría de los estratos aunque son mucho más abundantes en los arbóreos medio y alto.
En lo referente a la repoblación natural, se observó que para Dialium guianense, Billia rosea, Guatteria anomala y Dussia mexicana, entre 50 y 90 % de sus densidades se debió a individuos menores que 2 m de altura y diámetros menores que 4 cm. La contribución de dicha repoblación a la densidad global de su estrato representó 70 %. La repoblación natural es un fenómeno ampliamente reconocido en las selvas cálido húmedas y se favorece posiblemente por factores diversos, como la apertura natural de claros, condiciones ambientales ocasionalmente favorables y la capacidad propia de las especies para diseminarse y establecerse (Richards 1952, Martínez 1980, Brokaw 1985, Clark & Clark 1987, Hubbell & Foster 1987, Uhl et al. 1988, Martínez-Ramos et al. 1989, Whitmore 1989, Macario-Mendoza et al. 1992, 1995), aunque también es notable el hecho de que la mayoría de las especies del estrato arbóreo sobresaliente presenten bajas densidades en sus poblaciones (Sarukhán-Kermes 1968, Pérez-Jiménez & Sarukhán-Kermes 1970, Meave del Castillo 1983, Clark & Clark 1987, Hubbell & Foster 1987), posiblemente por su gran tolerancia y longevidad.
Por otra parte, cabe destacarse que las especies seleccionadas por los informantes lacandones como características de cada estrato en los rodales maduros de selva, fueron en todos los casos las mismas para las cuales se registraron los valores de importancia más altos en sus estratos respectivos. Con esto se ratifica la importancia que tiene el aprovechar los conocimientos tradicionales de los informantes locales sobre la flora y vegetación regional. En este mismo contexto, Levy-Tacher et al. (2002), Levy-Tacher & Golicher (2004), Levy-Tacher & Aguirre-Rivera (2005) y Durán-Fernández et al. (2016), con base en sus estudios sobre sucesión causada por aprovechamiento agrícola y las formas tradicionales alternativas de aprovechamiento persistente de la selva realizada por los lacandones de Lacanhá Chansayab reconocen la importancia del conocimiento ecológico tradicional en general, y etnobotánico en particular. La integración de los pobladores locales en el desarrollo de este tipo de estudios resulta crucial para asegurar la conservación de sus ecosistemas pues su participación los sensibiliza y revaloriza. Recientemente se ha implementado como una estrategia de conservación en la Selva Lacandona el pago por servicios ecosistémicos, particularmente con el 'Programa Especial Selva Lacandona (CONAFOR 2013) que ofrece beneficios (ca. 77 US$/ha/año) a propietarios y comunidades locales que preserven más que 25 ha de selva (Hernández-Ruedas et al. 2014).
La selva alta perennifolia de Nahá se encuentra en las partes altas marginales de la región Lacandona, lo que sugiere que está muy cerca de la zona de transición entre la tierra caliente y la templada (Breedlove 1981, Miranda 1993). Es una comunidad vegetal compleja, aún poco alterada por impacto de actividades extractivas comerciales, y estructuralmente similar al patrón general de otras variantes de selva alta (Miranda 1961, Sarukhán-Kermez 1968, Pérez-Jiménez & Sarukhán-Kermez 1970, Miranda 1975, 1976, Rzedowski 1978, Breedlove 1981, Meave del Castillo 1983, Bongers et al. 1988, Popma et al. 1988, Levy-Tacher 2000), pero presenta, como hemos denotado aquí, características distintivas, incluida la composición de especies y su distribución en los diferentes estratos.
Concluimos que el presente estudio permitió describir de forma detallada la composición y estructura de la selva alta perennifolia en la comunidad Lacandona de Nahá. El amplio conocimiento de los lacandones fue fundamental para el reconocimiento, tanto de los rodales de selva madura, como de los seis estratos bien definidos; lo que ratifica la importancia de incluir el conocimiento de los informantes locales en este tipo de estudios. Como en otras selvas de México existe un alto porcentaje de familias con una o pocas especies. Los atributos estructurales evaluados permitieron estimar la importancia de las especies en la comunidad y en los estratos correspondientes; éstos a su vez, presentaron composición florística y formas vitales distintivas. Terminalia amazonia fue la especie con el valor de importancia más alto, del estrato sobresaliente y de todos los demás estratos. Se resalta una relación inversa entre la densidad y el área basal, que al ser esta última un estimador de la biomasa, es el mejor indicador de dominancia en los sitios estudiados. La importancia de contar con análisis estructurales cuantitativos y detallados de los relictos de selva madura o bien conservada radica en que son primordiales como punto de referencia y fundamento para evaluar los efectos de los aprovechamientos y, en el futuro, diseñar las prácticas de restauración de las selvas altas perennifolias.

