




![Producción y viviparidad de nogal pecanero (Carya illinoinensis [Wangenh.] K. Koch) en relación con la humedad del suelo](/img/es/next.gif)





Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
versión On-line ISSN 2007-4018versión impresa ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.28 no.1 Chapingo ene./abr. 2022 Epub 24-Ene-2024
https://doi.org/10.5154/r.rchscfa.2021.07.045
Artículo científico
Anatomía de la madera de Magnolia rzedowskiana Vázquez, Domínguez & Pedraza, especie endémica de la Sierra Madre Oriental, México
1 Universidad Autónoma del Estado de Hidalgo, Centro de Investigaciones Biológicas. Ciudad del Conocimiento, carretera Pachuca-Tulancingo km 4.5. C. P. 42184. Mineral de la Reforma, Hidalgo, México.
2 Universidad Autónoma Chapingo, División de Ciencias Forestales, Laboratorio de Anatomía y Tecnología de la Madera. Carretera México-Texcoco km 38.5. C. P. 56230. Texcoco de Mora, Estado de México, México.
Introducción:
Magnolia rzedowskiana Vázquez, Domínguez & Pedraza es una especie arbórea endémica del bosque mesófilo de montaña de la Sierra Madre Oriental; sus poblaciones, poco densas y aisladas entre sí, se consideran en riesgo de extinción.
Objetivo:
Describir las características anatómicas microscópicas de la madera de M. rzedowskiana, para contribuir al conocimiento biológico de esta especie.
Materiales y métodos:
Las muestras de madera se obtuvieron por un método no destructivo, extrayendo dos núcleos de crecimiento de cinco árboles adultos en cada uno de los cuatro sitios de muestreo ubicados en las localidades Chilijapa y Zotictla en Hidalgo, La Yesca en Querétaro y ejido La Trinidad en San Luis Potosí. Los núcleos se seccionaron en submuestras, para elaborar preparaciones fijas a partir de cortes en sentido tangencial, transversal y radial. Las características se identificaron con microscopio óptico y se describieron de acuerdo con los criterios de la IAWA (International Association of Wood Anatomists).
Resultados y discusión:
Se diferenciaron zonas de crecimiento, porosidad difusa, poros solitarios y múltiples radiales en arreglo intermedio entre radial y tangencial; placa de perforación escaleriforme y fibras libriformes; parénquima axial paratraqueal y apotraqueal difuso y escaso; rayos biseriados combinados con triseriados, uniseriados y heterocelulares compuestos de células procumbentes con una hilera arriba y debajo de células cuadradas marginales con cuerpos de aceite. La madera se distingue por su mesomorfía y resistencia a la sequía.
Conclusión:
La descripción anatómica de M. rzedowskiana es un aporte al conocimiento biológico. En el sentido económico, la madera tiene potencial de uso para la producción de pulpa para papel.
Palabras clave: porosidad difusa; elementos de vaso; rayos; fibras libriformes; tilosis
Introduction:
Magnolia rzedowskiana Vázquez, Domínguez & Pedraza is a tree species endemic to the cloud forest of the Sierra Madre Oriental; its scanty (distributed) and isolated populations are considered at danger of extinction.
Objective:
To describe the microscopic anatomical characteristics of M. rzedowskiana wood to contribute to the biological knowledge of this species.
Materials and methods:
Wood samples were determined by a non-destructive method, extracting two growth cores from five adult trees in each of the four sampling sites located in Chilijapa and Zotictla in Hidalgo, La Yesca in Querétaro and ejido La Trinidad in San Luis Potosí. Cores were sectioned into subsamples to elaborate fixed preparations from tangential, transversal and radial cuts. Characteristics were identified using an optical microscopy and described according to IAWA (International Association of Wood Anatomists) criteria.
Results and discussion:
Growth zones, diffuse porosity, solitary and multiple radial pores in intermediate arrangement between radial and tangential; scalariform perforation plate and libriform fibers; diffuse and scanty paratracheal and apotracheal axial parenchyma; biseriate rays combined with triserial, uniseriate and heterocellular rays composed of procumbent cells with a row above and below of marginal square cells with oil bodies. The wood is distinguished by mesomorphism and resistance to drought.
Conclusion:
The anatomical description of M. rzedowskiana is a contribution to biological knowledge. From an economic point of view, this wood has potential use to produce pulp for paper.
Keywords: diffuse porosity; vessel elements; rays; libriform fibers; tylosis
Introducción
El estudio de la anatomía de la madera se aplica tanto a nivel tecnológico como en ciencia básica (Beeckman, 2016), ya que permite la elaboración de claves regionales para la identificación de especies (Araya-Montero & Moya-Roque, 2013), la reconstrucción de escenarios climáticos sobre la vida en las culturas prehispánicas y la generación de modelos para la predicción de las respuestas adaptativas de las plantas en las condiciones ambientales (Dickison, 2000). Los caracteres anatómicos en conjunto con los morfológicos pueden utilizarse para establecer relaciones o tendencias evolutivas, determinar familias, géneros y especies de plantas, y aclarar homologías en varias estructuras (Dickison, 2000; Wróblewska, 2015). Desde el enfoque de industrialización de la madera, el conocimiento de su anatomía permite hacer inferencias sobre las propiedades físicas y mecánicas, técnicas de procesamiento y uso potencial (Interián-Ku et al., 2011).
Los árboles y arbustos del género Magnolia L. se distribuyen ampliamente tanto en regiones templadas como en las tropicales, donde las especies son apreciadas por sus propiedades medicinales, ornamentales y maderables (Rico & Gutiérrez, 2019). El subgénero Magnolia se integra por 10 secciones del clado Tropical y Neotropical. Las secciones del clado Neotropical son Macrophylla, Manglietia, Magnolia, Oyama, Rytidospermum y Tuliparia, de acuerdo con la filogenia propuesta por Wang et al. (2020) con base en el genoma del cloroplasto. El árbol filogenético muestra relación directa entre el clado Macrophylla y el clado Tuliparia; este último incluye a la especie M. fraseri Walter y sus variedades endémicas del sureste de Estados Unidos (Wang et al., 2020). Las especies de la sección Macrophylla (M. alejandrae García-Morales & Iamonico, M. ashei Weatherby, M. dealbata Zuccarini, M. macrophylla Michaux, M. nuevoleonesis Vázquez & Domínguez-Yescas, Magnolia rzedowskiana Vázquez, Domínguez & Pedraza y M. vovidesii Vázquez, Domínguez-Yescas & Carvajal) se distribuyen en varias regiones entre el sureste de Estados Unidos y el sureste de México (Figlar & Nooteboom, 2004).
M. rzedowskiana (sección Macrophylla) es una especie descrita en el año 2015, considerada endémica y de distribución restringida; sus poblaciones sobreviven como relictos y están confinadas a la región central de la Sierra Madre Oriental, específicamente en la convergencia entre los estados de Querétaro (Sierra Gorda), San Luis Potosí (Sierra de Xilitla) e Hidalgo (Vázquez-García, Domínguez-Yescas, Pedraza-Ruiz, Sánchez-González, & Muñiz-Castro, 2015).
Los estudios sobre anatomía de la madera en especies de Magnolia son escasos, destaca el realizado en M. fraseri de la sección Tuliparia (InsideWood, 2004), el análisis del xilema secundario en M. schiedeana Schltdl. (Aguilar-Alcántara, Aguilar-Rodríguez, & Terrazas, 2014) y el trabajo sobre la plasticidad de los elementos de vaso ante la ocurrencia de sequías en M. schiedeana y M. vovidesii (Rodríguez-Ramírez, Vázquez-García, García-González, Alcántara-Ayala, & Luna-Vega, 2020). En este contexto, el objetivo del presente estudio fue describir las características anatómicas de la madera de las poblaciones de M. rzedowskiana en su área de distribución en la Sierra Madre Oriental en México, para aportar información biológica de la especie que actualmente se encuentra en riesgo de extinción por el grado de deterioro de su hábitat.
Materiales y métodos
Área de estudio
Las poblaciones analizadas de M. rzedowskiana se distribuyen en los estados de Hidalgo, Querétaro y San Luis Potosí, dentro de la Sierra Madre Oriental. La ubicación de estas poblaciones fue definida en un estudio previo sobre variación morfológica de la especie (Gutiérrez-Lozano, Sánchez-González, Vázquez-García, López-Mata, & Octavio-Aguilar, 2020). El tipo de vegetación con mayor cobertura en el área de estudio es bosque mesófilo de montaña, el clima predominante es templado húmedo (Instituto Nacional de Estadística y Geografía [INEGI], 2008), la precipitación total anual oscila entre 1 048 y 2 385 mm, y la temperatura promedio anual fluctúa entre 17.1 y 20.7 °C (Ruiz-Jiménez, Téllez-Valdés, & Luna-Vega, 2012).
Colecta de madera
Se eligieron cuatro sitios de muestreo ubicados en las localidades Chilijapa y Zotictla en Hidalgo, La Yesca en Querétaro y ejido La Trinidad en San Luis Potosí (Figura 1). En cada sitio de muestreo se seleccionaron cinco árboles adultos, sanos, de fuste recto. De cada árbol se obtuvieron dos núcleos de crecimiento con taladro de Pressler. Las muestras se extrajeron a 1.3 m de altura del tronco y los orificios se rellenaron con taquetes de madera previamente sumergidos en formol y se sellaron con resanador para madera. Adicionalmente, en cada sitio se registró la altitud y coordenadas geográficas para cada individuo (Cuadro 1).

Figura 1 Localización de los sitios de muestreo de las poblaciones de Magnolia rzedowskiana en la vertiente central de la Sierra Madre Oriental.
Cuadro 1 Localización geográfica y altitud de los sitios de muestreo de Magnolia rzedowskiana. Se incluyen datos del diámetro (a 1.30 m de altura) de los árboles utilizados para la extracción de las muestras de madera.
| Nombre del sitio | Coordenadas geográficas | Altitud (m) | Diámetro de los árboles (cm) | ||||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |||
| Ejido La Trinidad, Xilitla San Luis Potosí | 21° 24’ 0.62’’ N | 1 914 | 33.5 | 27.5 | 24.8 | 34 | 15 |
| 99° 03’ 54.8’’ O | |||||||
| La Yesca, Landa de Matamoros, Querétaro | 21° 13’ 2.9’’ N | 1 716 | 22.5 | 22.5 | 22 | 23 | 22 |
| 99° 07’ 52.6’’ O | |||||||
| Chilijapa, Tepehuacán de Guerrero, Hidalgo | 21° 00’ 40.4’’ N | 1 397 | 24 | 23 | 35 | 19 | 15.5 |
| 98° 52’ 24.3’’ O | |||||||
| Zotictla, Acaxochitlán, Hidalgo | 20° 13’ 20.4’’ N | 1 729 | 47 | 42 | 27 | 22.5 | 47.5 |
| 98° 09’ 40.6’’ O | |||||||
Preparaciones fijas
Los núcleos fueron seccionados en trozos de 1 cm (submuestra) y numerados de manera consecutiva en dirección corteza-médula. Se indujo un proceso de ablandamiento en agua destilada a 60 °C durante cinco días. Posteriormente, los núcleos se colocaron en una solución de glicerina, alcohol y agua en partes iguales (Borja-de la Rosa et al., 2010; Gärtner & Schweingruber, 2013). De cada submuestra se hicieron cortes de madera de 30 µm de grosor en los planos transversal, tangencial y radial con micrótomo de deslizamiento. Para estabilizar la estructura del tejido entre cortes se aplicó una solución de almidón de maíz (Gärtner et al., 2015). Los cortes se lavaron con una solución 3:7 de hipoclorito de sodio y agua destilada, se tiñeron con pardo de Bismarck durante dos horas y se deshidrataron con alcohol al 96 % y absoluto (100 %); posteriormente, el tejido se aclaró con xilol. El montaje se realizó con resina Entellan® (Borja de la Rosa et al., 2010; Navarro-Martínez, Borja-de la Rosa, & Machuca-Velasco, 2005).
Una pequeña parte de los núcleos de crecimiento se maceró de acuerdo con el método de Gärtner y Schweingruber (2013) para separar las células (fibras y vasos). Una vez disociado, el material se lavó y tiñó con pardo de Bismarck durante dos horas, se lavó el exceso de colorante y se hicieron preparaciones fijas para medir los elementos de vaso y fibras.
Descripción anatómica
La anatomía de la madera se describió con base en el documento “List of microscopic features for hardwood identification” (Wheeler, Bass, & Gasson, 1989), utilizando un microscopio óptico Motic. Los elementos constitutivos (rayos, elementos de vaso y fibras) se midieron y contaron en cada submuestra mediante imágenes digitales tomadas con una cámara Motic acoplada al microscopio y al software Motic Images 2000 (Motic®, 2000).
Análisis estadístico
Se estimaron las medidas de tendencia central (promedio aritmético) y de dispersión (desviación estándar, coeficiente de variación y rango) de los elementos constitutivos (rayos, elementos de vaso [EV] y fibras); además, se calcularon los índices de vulnerabilidad (IV) y mesomorfía (IM) (Carlquist, 1977) mediante las fórmulas siguientes:
También se calcularon los índices de calidad de pulpa para papel (ICP), a partir de los valores promedio de las fibras con respecto al diámetro (D), longitud (L), diámetro del lumen (I) y grosor de la pared celular (w). Las fórmulas para realizar los cálculos fueron las siguientes (Tamarit, 1996):
Resultados
La madera de M. rzedowskiana tiene bien definidas las zonas de crecimiento (Figura 2a), dado que están delimitadas por líneas de células de fibras aplanadas radialmente (Figura 2b), células de parénquima axial apotraqueal difuso (Figura 2c) y por el ensanchamiento de los rayos (Figura 2c).
La porosidad es difusa (Figura 2a); los vasos son solitarios y múltiples radiales de dos a nueve, en ocasiones arracimados de dos a siete; el arreglo es intermedio entre radial y tangencial; y el contorno predominante del vaso solitario es angular, esporádicamente se observan contornos circulares y ovalados (Figura 2d).

Figura 2 Corte transversal de la madera de Magnolia rzedowskiana: a) zona de crecimiento (ZC) definida por hileras de fibras y porosidad difusa (PD); b) fibras aplanadas en las zonas de crecimiento (Fa) y ensanchamiento del rayo (ER); c) células de parénquima axial apotraqueal difuso (PA); y d) detalles de las zonas de crecimiento y poros (ZT = zona de transición, FMTa = fibras de madera en zonas de transición, ER = ensanchamiento del rayo, CVA = contorno angular del elemento de vaso, EVS = elemento de vaso solitario y EVM = elementos de vaso múltiples).
La placa de perforación en los EV es escaleriforme (con una a 15 barras) y simple (Figura 3a), con puntuaciones intervasculares escaleriformes (Figura 3b) y radiovasculares horizontales escaleriformes (Figura 4a) y con engrosamientos helicoidales en la base del vaso (Figura 4b). Los EV son cilíndricos, alargados, de varios tamaños con y sin colas pequeñas en ambos extremos (Figura 5a); la longitud promedio es de 785.55 ± 151.74 µm y diámetro de muy pequeño a mediano (≤50 µm, 50-100 µm, 100-200 µm) con promedio de 58.77 ± 10.37 µm. En la sección transversal se contabilizaron 60.03 ± 17.06 vasos∙mm-2. El Cuadro 2 muestra las medidas de tendencia central y de dispersión estimadas. Por otra parte, se observaron EV con depósitos de aceite hacia la zona de la médula (Figura 6a) y en raras ocasiones con presencia de tilosis (Figura 6c).

Figura 3 Elementos de vaso de Magnolia rzedowskiana: a) Placa de perforación escaleriforme (PPE) y punteadura de la fibra areolada elíptica (PF) en la sección radial; b) punteadura escaleriforme (PE) en la sección tangencial.

Figura 4 Sección radial de Magnolia rzedowskiana: a) Punteadura radio vascular-escaleriforme (PVR) y b) engrosamiento helicoidal en elemento del vaso (EEV).
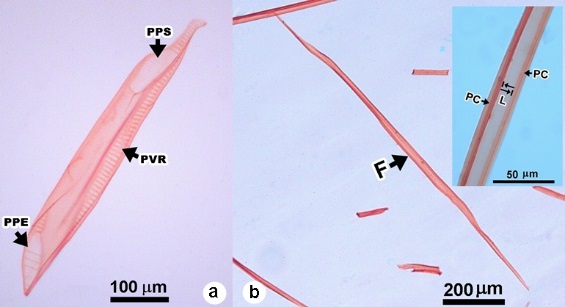
Figura 5 a) Elementos de vaso de Magnolia rzedowskiana (PPS = placa de perforación simple, PVR = punteadura radio vascular-escaleriforme y PPE = placa de perforación escaleriforme). b) Fibra libiforme (F) y detalle de la fibra (PC = pared celular y L = lumen).

Figura 6 Elementos de vaso y rayo de Magnolia rzedowskiana: a) Depósito de aceite (Ca) y tilosis (T) en la sección transversal, b) depósito de aceite (Ca) en sección radial, y c) tilosis (T).
Las fibras son medianas (900 a 1 600 µm) de tipo libriforme (Figura 5b) y poseen punteaduras areoladas elípticas. Algunas fibras tienen septas (Figura 7a) con engrosamiento helicoidal de longitud promedio de 1 333.07 ± 197.69 µm y diámetro promedio de 26.33 ± 4.35 µm. El diámetro promedio del lumen es de 11.79 ± 2.89 µm y el grosor promedio de la pared celular es 14.55 ± 3.31 µm (Cuadro 2).
Los rayos de una a tres células de ancho, en su mayoría, son de dos series en combinación con rayos de tres y una serie (Figura 7b) y en ocasiones con rayos agregados (Figura 7c). El número de rayos fue 6.13 ± 1.2 por mm lineal con altura de 641.57 ± 150.90 µm y ancho de 54.67 ± 14.86 µm (Cuadro 2). En la sección radial, los rayos son heterocelulares (Figura 8a) compuestos de células procumbentes, con una hilera de células marginales cuadradas arriba y debajo. La estructura estratificada de los rayos es baja y, debido a su variación de altura, no se forman filas de rayos horizontales uniformes (Figura 8b); en algunos fue posible observar inclusiones de sílice (compuestas de dióxido de silicio) de forma esférica (Figura 8c) y depósitos de aceite (Figura 6b).

Figura 7 Fibras (FS) y rayos (R) de la madera de Magnolia rzedowskiana: a) sección tangencial; b) tipos de rayos (RU = rayos de una serie, RB = rayos de dos series, RT = rayos de tres series); c) rayo agregado (RA).

Figura 8 a) Rayo heterocelular (células erectas [CE] y células procumbentes [CP]) en la sección radial de Magnolia rzedowskiana; b) estratificación baja en los rayos; c) inclusiones de sílice (CS); y d) parénquima axial paratraquial (PP) y apotraqueal (PA).
Cuadro 2 Valores promedio y de dispersión de las células que constituyen la madera de Magnolia rzedowskiana.
| Elemento constitutivo | Parámetro estadístico | n | Media | DE | CV (%) | Rango | Máximo | Mínimo | Clasificación (Tortorelli, 1956) |
|---|---|---|---|---|---|---|---|---|---|
| Elemento de vaso | Diámetro (mm) | 4 070 | 58.77 | 10.37 | 17.52 | 74.8 | 105.66 | 30.86 | Pequeños (<100 mm) |
| Longitud (mm) | 4 070 | 785.55 | 151.74 | 19.32 | 1 152.53 | 1 384.08 | 231.55 | Medianos (350-800 mm) | |
| Vasos∙mm-2 | 411 | 60.03 | 17.06 | 28.42 | 142 | 167 | 25 | Extremadamente numerosos(>40) | |
| Fibras | Longitud (mm) | 4 070 | 1337.07 | 197.69 | 14.79 | 1 486.06 | 2 090.46 | 604.34 | Medianas (900-1 600 mm) |
| Diámetro (mm) | 4 070 | 26.33 | 4.35 | 16.51 | 37.45 | 48.3 | 10.85 | Medianas (25-40 mm) | |
| Grosor pared celular (mm) | 4 070 | 14.55 | 3.31 | 22.72 | 34.52 | 36.73 | 2.21 | Gruesa (lumen de 1/2 a 1/3 del diámetro total) | |
| Lumen (mm) | 4 070 | 11.79 | 2.89 | 24.5 | 24.82 | 26.2 | 1.38 | - | |
| Rayos | Altura (mm) | 2 207 | 641.57 | 150.9 | 23.52 | 1 394.60 | 1 619.30 | 224.7 | Medianos (500-800 mm) |
| Ancho (mm) | 2 207 | 54.67 | 14.86 | 27.17 | 101.5 | 124.4 | 22.9 | Medianos (50-100 mm) | |
| Rayos∙mm-2 | 410 | 6.13 | 1.2 | 19.51 | 8 | 11 | 3 | Poco numerosos (5-7) |
n: tamaño de la muestra, DE: desviación estándar, CV: coeficiente de variación.
La madera contiene parénquima axial paratraqueal (PP) y parénquima axial apotraqueal (PA) (Figura 8d). Las células del PP son escasas puesto que no cubren totalmente el contorno del EV. Las células del PA también son escasas y están dispersas a lo ancho de las zonas de crecimiento sin formar bandas o hileras; algunas células de PA se intercalan entre las fibras de transición en las zonas de crecimiento (Figura 2c).
Con respecto a la calidad de la madera, los valores estimados de los índices de vulnerabilidad, mesomorfía y calidad de pulpa para papel se muestran en el Cuadro 3.
Cuadro 3 Índices utilizados para estimar las características anatómicas de la madera de Magnolia rzedowskiana.
| Índice | Valor estimado | Rango | Clasificación |
|---|---|---|---|
| Índice de vulnerabilidad | 0.98 | <1 | Resistente a sequía (Carlquist, 1977) |
| Índice de mesomorfía | 769.1 | <200 | Xilema mesofítico (Carlquist, 1977) |
| Coeficiente o índice de rigidez | 1.1 | >0.70 | Muy gruesa (Villaseñor & Rutiaga, 2000) |
| Coeficiente de flexibilidad | 0.45 | 0.30 – 0.50 | Gruesa; las fibras se colapsan muy poco; poca superficie de contacto; poca unión fibra con fibra (Villaseñor & Rutiaga, 2000) |
| Coeficiente de Peteri o índice de esbeltez | 91.91 | >75 | El material fibroso tiende a plastificarse y tiene buena superficie de contacto; es decir, tiene buena adherencia entre fibras (Tamarit, 1996; Zaragoza-Hernández, Borja-de la Rosa, Zamudio-Sanchez, Ordoñez-Candelaria, & Bárcenas-Pazos, 2014) |
| Relación de Runkel | 2.47 | Grado I, <0.25 | Excelente (Tamarit, 1996; Villaseñor & Rutiaga, 2000) |
Discusión
La madera de M. rzedowskiana tiene caracteres semejantes a los de M. fordiana (Oliv.) Hu, M. fraseri, M. grandiflora L., M. obovata Thunb., M. schiedeana, M. virginiana L., todas del subgénero Magnolia (Figlar & Nooteboom, 2004; Wang et al., 2020). Las características comunes en estas especies son: zonas de crecimiento definidas, porosidad difusa, contorno angular del vaso solitario, placas de perforación escaleriforme (≤10 barras), punteadura intervascular escaleriforme, diámetro tangencial de los vasos medianos (100 a 200 µm), longitud promedio de los vasos medianos (350 a 800 µm), fibras no septadas, rayos de una a tres células (uniseriados, biseriados y triseriados) y rayos heterocelulares compuestos de células procumbentes con una hilera de células marginales cuadradas arriba y debajo (InsideWood, 2004).
La madera temprana en especies de porosidad difusa constituye casi todo el anillo de crecimiento que se formó durante varios meses, mientras que la madera tardía está representada por zonas de crecimiento muy estrechas con fibras aplanadas (Schweingruber, 2007). En M. rzedowskiana se observaron de dos a tres hileras de fibras aplanadas que constituyen la transición entre zonas de crecimiento, las cuales se encuentran bien definidas.
La porosidad difusa en M. rzedowskiana es una característica común en otras especies de magnolias (InsideWood, 2004; Wróblewska, 2015). Este tipo de porosidad se caracteriza por el diámetro de los elementos de vasos, que es más o menos semejante en todo el anillo de crecimiento (Wheeler et al., 1989). En especies de porosidad difusa, el incremento de diámetro en el tallo inicia después de la aparición de las hojas, y la producción de vasos es continua y de menor intensidad durante la anualidad, mientras que, en especies de porosidad circular y semicircular, los vasos funcionales se producen de forman intensiva en un periodo corto (Takahashi, Okada, & Nobuchi, 2013). El tamaño y densidad de los vasos en especies de porosidad difusa está relacionado estrechamente con el metabolismo de los árboles y revelan las consecuencias de las limitaciones climáticas en el crecimiento de los individuos; en condiciones de sequía, el crecimiento radial es reducido, el área del lumen de los vasos es menor y la densidad de vasos aumenta como un mecanismo de adaptación para la seguridad hidraúlica y para evitar el riesgo de embolia y cavitación (Islam, Rahman, & Bräuning, 2018).
En M. rzedowskiana, los EV son solitarios, aunque también se encontraron múltiples radiales y en racimos en menor proporción, característica que comparte con M. ovata y M. grandiflora (Ruffinato & Crivellaro, 2019). Los vasos múltiples radiales ofrecen una alternativa para que el sistema conductor produzca nuevos elementos, que pueden asumir la función de los vasos formados previamente sin alterar las vías conductoras (Carlquist, 2001).
La placa de perforación escaleriforme en vasos con 10 a 20 barras es un rasgo común entre M. rzedowskiana, M. fraseri, M. grandiflora, M. obovata, M. virginiana (InsideWood, 2004) y M. wilsonii (Finet & Gagnepain) Rehder; en esta última especie, algunas barras de la placa escaleriforme se bifurcan (Wróblewska, 2015), peculiaridad que también se observó en M. rzedowskiana. Es importante destacar que rara vez se observaron placas de perforación simple; de acuerdo con Canright (1955), la presencia de este tipo de placas en algunos EV es común en especies de clima templado. Las especies de Magnolia se caracterizan por sus punteaduras intervasculares de tipo escaleriforme (Carlquist, 1988; Takhtajan, 2009), las cuales son comunes en especies con vasos angulares (poligonales), también ocurren en la interfaz rayo vascular, aunque no son comunes en las dicotiledóneas (Carlquist, 2001). El engrosamiento helicoidal en la base de los EV en M. rzedowskiana también se encuentra en M. fordiana, M. grandiflora, M. schiedeana y M. virginiana (InsideWood, 2004).
La presencia de tilosis es rara, pero está presente en M. rzedowskiana, M. cordata Michaux, M. pterocarpa Roxburgh y M. tripetala L. (Canright, 1955). La formación de tilosis se debe a la pérdida de presión del agua en los vasos; su origen parece ser esencialmente fisiológico pues no se ha identificado algún patrón filogenético de distribución de tílides en especies arbóreas (Carlquist, 2001).
Las fibras en M. rzedowskiana son libriformes con puntuaciones areoladas elípticas y algunas fibras septadas, diferentes a las que presentan M. fordiana, M. fraseri, M. grandiflora, M. obovata, M. schiedeana y M. virginiana con punteaduras simples y sin septas (InsideWood, 2004). Al respecto, Metcalfe (1950) menciona que las fibras de algunas especies de Magnolia tienen septas y Takhtajan (2009) refiere que las fibras poseen punteaduras rebordeadas.
De acuerdo con Takhtajan (2009), en la madera de las magnolias, el parénquima axial es de tipo apotraqueal difuso y en la mayoría de las especies su distribución es terminal, lo cual puede ser atribuido directamente a cuestiones fisiológicas como la caída anual de las hojas (excepto en especies tropicales no caducifolias). Słupianek, Dolzblasz, y Sokołowska (2021) indican que, en especies de árboles de angiospermas, la fracción de parénquima axial es muy variable (rango ≤1 hasta ≥30 %) y que el género Magnolia se caracteriza por un parénquima axial escaso; sin embargo, en algunas especies también se han encontrado bandas de parénquima al comienzo del anillo de crecimiento (Canright, 1955). En el caso de M. rzedowskiana se encontraron células de parénquima axial apotraqueal dispersas sin formar hileras. Carlquist (2001) menciona que de los tipos de parénquima, el difuso es el más primitivo, puede ser escaso y se distribuye a manera de hebras axiales esparcidas al azar dentro del xilema secundario.
Las células con contenidos de aceites, características de Magnolia (Metcalfe, 1950), se observaron en los vasos y rayos de M. rzedowskiana, pero en M. fordiana y M. schiedeana, al parecer, solo se presentan en los rayos (InsideWood, 2004).
Por otra parte, los índices de vulnerabilidad y de mesomorfía revelan que M. rzedowskiana tiene un sistema eficaz de conducción del agua que favorece la resistencia a condiciones de estrés hídrico (Carlquist, 1977). De acuerdo con la relación de Runkel, la calidad de las fibras de M. rzedowskiana es excelente para obtener papel, con grado de calidad I, por lo cual la madera tiene potencial para la producción de papel resistente, transparente y con buena adhesión de fibra a fibra (Tamarit, 1996).
Conclusiones
Las características anatómicas de la madera de M. rzedowskiana que destacan con respecto a otras especies de Magnolia son: diámetro de elementos de vaso más angostos, ocurrencia ocasional de placas de perforación simples, fibras septadas, tilosis y algunos rayos agregados; si bien, no son caracteres exclusivos y predominantes en la especie, sí son distintivos. Esta descripción contribuye al conocimiento de la especie para el desarrollo de investigaciones enfocadas en el manejo y conservación de las poblaciones, germinación de semillas, producción de plántulas en vivero y tecnología de la madera. Los resultados son importantes desde el punto de vista utilitario, ya que indican que la madera de esta especie puede tener relevancia económica a nivel local. Como las poblaciones de M. rzedowskiana son actualmente muy pequeñas y dispersas, se recomienda el desarrollo de programas de conservación en su área de distribución.
References
Aguilar-Alcántara, M., Aguilar-Rodríguez, S., & Terrazas, T. (2014). Anatomía de la madera de doce especies de un bosque mesófilo de montaña de Tamaulipas, México. Madera y Bosques, 20(3), 69-86. Retrieved from http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S1405-04712014000300007&lng=es&tlng=es [ Links ]
Araya-Montero, M., & Moya-Roque, R. (2013). Claves de identificación macroscópica de la madera de 110 especies del Caribe Norte de Costa Rica. Revista Forestal Mesoamericana Kurú, 10(24), 34-52. doi: 10.18845/rfmk.v10i24.1322 [ Links ]
Beeckman, H. (2016). Wood anatomy and trait-based ecology. IAWA Journal, 37(2), 127-151. doi: 10.1163/22941932-20160127 [ Links ]
Borja-de la Rosa, A., Machuca, R., Salinas, M. F., Lozada, D. A., López, M. F., & Alcantar, A. Q. (2010). Caracterización tecnológica de la madera de Juniperus flaccida var. poblana Martínez. Revista Chapingo Serie Ciencias Forestales y del Ambiente, 16(2), 261-280. doi: 10.5154/r.rchscfa.2010.09.083 [ Links ]
Canright, J. E. (1955). The comparative morphology and relationships of the Magnoliacea-IV. Wood and nodal anatomy. Journal of the Arnold Arboretum, 36(2), 119-140. Retrieved from http://www.jstor.org/stable/43790865 [ Links ]
Carlquist, S. (1977). Ecological factors in wood evolution: a floristic approach. American Journal of Botany, 64(7), 887-896. doi: 10.2307/2442382 [ Links ]
Carlquist, S. (1988). Comparative wood anatomy. Systematic, ecological and evolutionary aspects of dicotyledon wood. Berlin, Heidelberg: Springer-Verlag. doi: 10.1007/978-3-662-21714-6 [ Links ]
Carlquist, S. (2001). Comparative wood anatomy: systematic, ecological, and evolutionary aspects of dicotyledon wood (2nd. ed.). Berlin, Heidelberg: Springer-Verlag . doi: 10.1007/978-3-662-04578-7 [ Links ]
Dickison, W. C. (2000). Integrated plant anatomy. Academic Press. doi: 10.1016/B978-0-12-215170-5.X5000-6 [ Links ]
Figlar, R. B., & Nooteboom, H. P. (2004). Notes on Magnoliaceae IV. Blumea-Biodiversity, Evolution and Biogeography of Plants, 49(1), 87-100. doi: 10.3767/000651904X486214 [ Links ]
Gärtner, H., Cherubini, P., Fonti, P., Von Arx, G., Schneider, L., Nievergelt, D., & Büntgen, U. (2015). A technical perspective in modern tree-ring research - How to overcome dendroecological and wood anatomical challenges. Journal of Visualized Experiments, 97, e52337. doi: 10.3791/52337 [ Links ]
Gärtner, H., & Schweingruber, F. H. (2013). Microscopic preparation techniques for plant stem analysis. Remagen, Germany: Verlag Dr. Kessel. Retrieved from https://www.researchgate.net/publication/253341899_Microscopic_Preparation_Techniques_for_Plant_Stem_Analysis [ Links ]
Gutiérrez-Lozano, M., Sánchez-González, A., Vázquez-García, J. A., López-Mata, L., & Octavio-Aguilar, P. (2020). Diferenciación morfológica poblacional de Magnolia rzedowskiana (Magnoliaceae): especie endémica en peligro de extinción de la sierra Madre Oriental, México. Revista Mexicana de Biodiversidad, 91, 1-14. doi: 10.22201/ib.20078706e.2020.91.3101 [ Links ]
InsideWood. (2004). The InsideWood Database. Retrieved November, 2020, from http://insidewood.lib.ncsu.edu/search [ Links ]
Instituto Nacional de Estadística y Geografía (INEGI). (2008). Carta Climatológica - Unidades Climaticas. Escala 1:1 000 000. Aguascalientes, México: Author. Retrieved from https://www.inegi.org.mx/app/biblioteca/ficha.html?upc=702825267568 [ Links ]
Interián-Ku, V. M., Borja-de la Rosa, M. A., Valdez-Hernández, J. I., García-Moya, E., Romero-Manzanares, A., & Vaquera-Huerta, H. (2011). Características anatómicas y propiedades físicas de la madera de Caesalpinia gaumeri Greenm en Dzan, Yucatán. Madera y Bosques, 17(1), 23-36. Retrieved from http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S1405-04712011000100002&lng=es&tlng=es [ Links ]
Islam, M., Rahman, M., & Bräuning, A. (2018). Xylem anatomical responses of diffuse porous Chukrasia tabularis to climate in a South Asian moist tropical forest. Forest Ecology and Management, 412, 9-20. doi: 10.1016/j.foreco.2018.01.035 [ Links ]
Metcalfe, C. R., & Chalk, L. (1950). Anatomy of the Dicotyledons: leaves, stem, and wood, in relation to taxonomy, with notes on economic uses. Oxford, Great Britain: Clarendon Press. [ Links ]
Motic® (2000). Motic images 2000 (version 1.3). China: Motic China Group CO., LTD. [ Links ]
Navarro-Martínez, J., Borja-de la Rosa, A., & Machuca-Velasco, R. (2005). Características tecnológicas de la madera de palo morado (Peltogyne mexicana Martínez) de Tierra Colorada, Guerrero, México. Revista Chapingo Serie Ciencias Forestales y del Ambiente, 11(1), 73-82. Retrieved from https://www.redalyc.org/articulo.oa?id=62911110 [ Links ]
Rico, Y., & Gutiérrez, B. B. A. (2019). Species delimitation and genetic structure of two endemic Magnolia species (section Magnolia; Magnoliaceae) in Mexico. Genetica, 147(1), 57-68. doi: 10.1007/s10709-019-00052-8 [ Links ]
Rodríguez-Ramírez, E. C., Vázquez-García, J. A., García-González, I., Alcántara-Ayala, O., & Luna-Vega, I. (2020). Drought effects on the plasticity in vessel traits of two endemic Magnolia species in the tropical montane cloud forests of eastern Mexico. Journal of Plant, 13(3), 331-340. doi: 10.1093/jpe/rtaa019 [ Links ]
Ruffinatto, F., & Crivellaro, A. (2019). Atlas of macroscopic wood identification: With a special focus on timbers used in Europe and CITES-listed species. Springer. doi: 10.1007/978-3-030-23566-6 [ Links ]
Ruiz-Jiménez, C. A., Téllez-Valdés, O., & Luna-Vega, I. (2012). Clasificación de los bosques mesófilos de montaña de México: afinidades de la flora. Revista Mexicana de Biodiversidad, 83(4), 1110-1144. Retrieved from http://www.scielo.org.mx/pdf/rmbiodiv/v83n4/v83n4a19.pdf [ Links ]
Schweingruber, F. H. (2007). Wood structure and environment. Springer. doi: 10.1007/978-3-540-48548-3 [ Links ]
Słupianek, A., Dolzblasz, A., & Sokołowska, K. (2021). Xylem parenchyma-role and relevance in wood functioning in trees. Plants, 10(6), 1247. doi: 10.3390/plants10061247 [ Links ]
Takahashi, S., Okada, N., & Nobuchi, T. (2013). Relationship between the timing of vessel formation and leaf phenology in ten ring-porous and diffuse-porous deciduous tree species. Ecological Research, 28(4), 615-624. doi: 10.1007/s11284-013-1053-x [ Links ]
Takhtajan, A. (2009). Flowering plants. Dordrecht: Springer. doi: 10.1007/978-1-4020-9609-9 [ Links ]
Tamarit, U. J. C. (1996). Determinación de los índices de calidad de pulpa para papel de 132 maderas latifoliadas. Madera y Bosques, 2(2), 29-41. doi: 10.21829/myb.1996.221384 [ Links ]
Tortorelli, A. (1956). Maderas y bosques de Argentina. Buenos Aires, Argentina: ACME. [ Links ]
Vázquez-García, J. A., Domínguez-Yescas, R., Pedraza-Ruiz, R., Sánchez-González, A., & Muñiz-Castro, M. Á. (2015). Magnolia rzedowskiana (Magnoliaceae), una especie nueva de la sección Macrophylla de la parte central de la Sierra Madre Oriental, México. Acta Botánica Mexicana, 112, 19-36. doi: 10.21829/abm112.2015.1086 [ Links ]
Villaseñor, A. J. C., & Rutiaga, Q. J. G. (2000). La madera de Casuarina equisetifolia L., química e índices de calidad de pulpa. Madera y Bosques, 6(1), 29-40. doi: 10.21829/myb.2000.611340 [ Links ]
Wang, Y. B., Liu, B. B., Nie, Z. L., Chen, H. F., Chen, F. J., Figlar, R. B., & Wen, J. (2020). Major clades and a revised classification of Magnolia and Magnoliaceae based on whole plastid genome sequences via genome skimming. Journal of Systematics and Evolution, 58(5), 673-695. doi: 10.1111/jse.12588 [ Links ]
Wheeler, E. A., Bass, P., & Gasson, P. E. (1989). IAWA list of microscopic features for hardwood identification. IAWA Bulletin n.s., 10(3), 219-332. Retrieved from https://www.iawa-website.org/uploads/soft/Abstracts/IAWA%20list%20of%20microscopic%20features%20for%20hardwood%20identification.pdf [ Links ]
Wróblewska, M. M. (2015). The progressive and ancestral traits of the secondary xylem within Magnolia clad - the early diverging lineage of flowering plants. Acta Societatis Botanicorum Poloniae, 84(1), 87-96. doi: 10.5586/asbp.2014.028 [ Links ]
Zaragoza-Hernández, I., Borja-de la Rosa, A., Zamudio-Sánchez, F. J., Ordóñez-Candelaria, V. R., & Bárcenas-Pazos, G. M. (2014). Anatomía del culmo de bambú (Guadua aculeata Rupr.) de la región nororiental del estado de Puebla, México. Madera y Bosques, 20(3), 87-96. doi: 10.21829/myb.2014.203154 [ Links ]
Recibido: 11 de Julio de 2021; Aprobado: 03 de Diciembre de 2021
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
