



![Modelado espacial histórico y actual del oyamel (Abies religiosa [Kunth] Schltdl. & Cham.) en la Faja Volcánica Transmexicana](/img/es/prev.gif)






Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
versión On-line ISSN 2007-4018versión impresa ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.25 no.2 Chapingo may./ago. 2019 Epub 19-Feb-2021
https://doi.org/10.5154/r.rchscfa.2018.09.066
Artículo científico
Influencia del método de árboles padres en la diversidad de la regeneración de bosques mixtos de Durango, México
1Instituto Tecnológico de El Salto. Calle Tecnológico col. La Forestal. C. P. 34942. El Salto, Durango, México.
Introducción:
El manejador forestal necesita conocer el impacto que las cortas de regeneración causan en la diversidad de especies arbóreas, para poder conservarla.
Objetivo.
Evaluar los cambios en riqueza y diversidad de especies arbóreas después de aplicar el método de árboles padres en seis localidades.
Materiales y métodos.
Las localidades Ciénega Grande, Las Ciénegas, La Colmena, El Venado, Los Bajíos y Pino Gordo del ejido Pueblo Nuevo, Durango, se dividieron en tres parcelas denominadas: a) regeneración en árboles padres (RAP); b) regeneración en arbolado adulto (RAA); y c) arbolado adulto (AA). Partiendo del centro de cada parcela de RAP, y siguiendo los puntos cardinales y subcardinales, se ubicaron ocho sitios de 25 m2 y 0.1 ha, para registrar las especies de la regeneración y las del arbolado adulto, respectivamente. Los indicadores de riqueza de especies, Shannon-Wiener, Simpson, Pielou y Sørensen se estimaron para caracterizar las parcelas por localidad y se compararon con pruebas de t y análisis de rarefacción.
Resultados y Discusión.
Se registraron 12 especies presentes en el área de estudio, de las cuales 11 se encuentran como renuevo de árboles padres. La riqueza de especies en las parcelas de RAP con respecto a la de AA fue igual en las localidades Ciénega Grande, El Venado y La Colmena, y superior en Las Ciénegas y Los Bajíos. Los índices de Shannon-Wiener y Simpson fueron superiores en Pino Gordo, Los Bajíos y El Venado. El índice de Sørensen varía de 0.67 a 1.0, indicando alta semejanza de especies entre parcelas.
Conclusiones.
El método de árboles padres garantiza la continuidad de la diversidad de especies arbóreas.
Palabras clave: Riqueza de especies; Shannon-Wiener; Simpson; Pielou; Sørensen; rarefacción
Introduction:
Forest managers need to know the impact of regeneration cuts on tree species diversity in order to conserve it.
Objective:
To evaluate changes in tree species richness and diversity after applying the seed tree method in six communities.
Materials and methods:
The communities of Ciénega Grande, Las Ciénegas, La Colmena, El Venado, Los Bajíos and Pino Gordo of the Pueblo Nuevo ejido, Durango, were divided into three plots called: a) regeneration in seed trees cuttings (RST); b) regeneration in adult trees (RAT); and c) adult trees (AT). Starting from the center of each RST plot, and following the cardinal and subcardinal points, eight sites of 25 m2 and 0.1 ha were located to record the regeneration and adult tree species, respectively. Species richness indicators (Shannon-Wiener, Simpson, Pielou and Sørensen) were estimated to characterize the plots by community and were compared with t-tests and rarefaction analysis.
Results and discussion:
Twelve species were recorded in the study area, of which 11 are found as a renewal of seed trees. Species richness in the RST plots with respect to that of AT was equal in the Ciénega Grande, El Venado and La Colmena communities, and higher in Las Ciénegas and Los Bajíos. The Shannon-Wiener and Simpson indices were higher in Pino Gordo, Los Bajíos and El Venado. The Sørensen index ranges from 0.67 to 1.0, indicating high species similarity among plots.
Conclusions.
The seed tree method in a mixed forest ensures the continuity of tree species diversity.
Keywords: Species richness; Shannon-Wiener; Simpson; Pielou; Sørensen; rarefaction
Introducción
Una de las premisas en la ejecución de programas de manejo forestal es la conservación de la diversidad de las especies arbóreas, especialmente cuando el objetivo principal es proveer el hábitat de recursos para los organismos presentes en el bosque (Jayakumar & Nair, 2013). No obstante, la conservación de la diversidad se pone en riesgo cuando se pretende favorecer el aprovechamiento de las especies maderables de rápido crecimiento, bajo sistemas de manejo intensivo. Los sistemas intensivos de manejo regular buscan maximizar la producción de madera, disminuyendo la presencia de especies arbóreas, para favorecer el establecimiento de especies deseables de rápido crecimiento. En contraste, los sistemas de bosques irregulares buscan optimizar la producción de madera manteniendo el equilibrio dinámico del ecosistema, bajo el principio de la conservación de la estructura y composición de especies. La simplificación de la diversidad arbórea reduce las funciones del sistema, mientras que su complejidad las incrementa, mejorando el nivel de productividad (Castellanos-Bolaños, Treviño-Garza, Aguirre-Calderón, Jiménez-Pérez, & Velázquez-Martínez, 2010; Ishii, Tanabe, & Hiura, 2004).
La conservación de la composición de las especies arbóreas depende de su potencial de regeneración, considerando que el potencial es la habilidad para completar cada etapa de desarrollo, y que la regeneración es un proceso biológico estratégico que garantiza la presencia de las especies en una comunidad (Khumbongmayum, Khan, & Tripathi, 2005). En el establecimiento de la regeneración natural en bosques mixtos, es importante evaluar los atributos poblacionales y de crecimiento de las plantas, para detectar los posibles cambios generados en la composición y diversidad dentro del rango de distribución geográfica de cada especie (Grubb, 1977; Malik & Bhatt, 2016). La presencia de bosques mixtos, caracterizados por una diversidad alta de especies y diferenciación estructural, demanda conocer la respuesta del arbolado a la aplicación de tratamientos silvícolas, en especial el destinado a establecer la regeneración, para definir las prácticas que conlleven a la sustentabilidad del manejo forestal (Corral, Aguirre, Jiménez, & Corral, 2005; Gadow, Sánchez, & Aguirre, 2004; Solís et al., 2006). Los métodos de regeneración modifican las condiciones ambientales y la disponibilidad de los recursos, así como la capacidad de reproducción o regeneración de las especies. La aplicación de estos métodos garantiza la presencia de las especies vegetales bajo diversas condiciones ambientales, para conservar la composición (Khumbongmayum et al., 2005: Smith, Larson, Kelty, & Ashton, 1997).
Los bosques del suroeste de Durango se caracterizan por ser mixtos con la presencia de especies de Pinus y Quercus (Medrano-Meraz, Hernández, Corral-Rivas, & Nájera-Luna, 2017); sin embargo, aunque se pretende conservar la diversidad primaria, se aplican sistemas de manejo regular que favorecen la regulación de la composición de especies, a través de la aplicación de la corta de árboles padres. Este tratamiento básico del sistema de manejo regular puede promover cambios en la presencia y diversidad de especies. Poco se ha documentado sobre la respuesta de dicho tratamiento, en términos del establecimiento de especies diferentes a las de los árboles padres; en este sentido, el objetivo del presente estudio fue evaluar el efecto de la aplicación de esta corta de regeneración sobre la diversidad de especies arbóreas presentes en el sotobosque.
Materiales y métodos
Ubicación y descripción del área de estudio
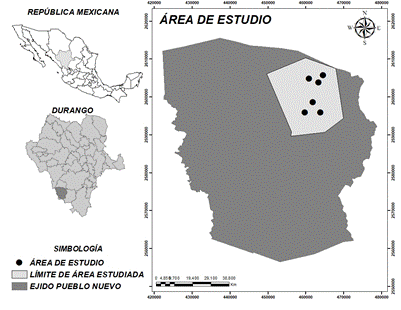
El estudio se llevó a cabo en el ejido Pueblo Nuevo, ubicado en el macizo montañoso conocido como Sierra Madre Occidental en la región de El Salto, al suroeste del estado de Durango, México (Figura 1). El clima es semifrío subhúmedo con lluvias en verano y temperatura media anual entre 5 y 12 °C; la temperatura del mes más frío varía entre 3 y 18 °C con precipitación durante el invierno entre 5 y 10.2 % (García, 1981). De acuerdo con la Carta Edafológica Serie II, el suelo del área de estudio se clasifica como Regosol (Instituto Nacional de Estadística y Geografía [INEGI], 2010), mientras que el tipo de vegetación es de pino-encino.
Muestreo
Se seleccionaron seis localidades donde se aplicó el método de regeneración de árboles padres en un periodo anterior a 10 años y sin tratamiento silvícola posterior, bajo las siguientes denominaciones: Las Ciénegas, Ciénega Grande, La Colmena, El Venado, Los Bajíos y Pino Gordo. Cada localidad se dividió en tres parcelas identificadas como: a) regeneración en corta de árboles padres (RAP), b) regeneración bajo arbolado adulto (RAA) y c) arbolado adulto (AA). Las parcelas de RAP se caracterizan por presentar especies arbóreas que se establecieron en el sotobosque después de haber aplicado el tratamiento de regeneración de árboles padres; estos árboles son individuos de Pinus durangensis Martínez y P. cooperi C. E. Blanco, especies de mayor importancia económica. A su vez, las parcelas de RAA presentan regeneración avanzada, establecida bajo la cobertura del arbolado adulto adyacente a las parcelas donde se aplicó la corta de árboles padres. Finalmente, las parcelas AA están compuestas por el arbolado mayor de 7.5 cm de diámetro a la altura de 1.30 m del suelo y que, al igual que la RAA, se encuentran adyacentes a la parcela donde se aplicó la corta de árboles padres.
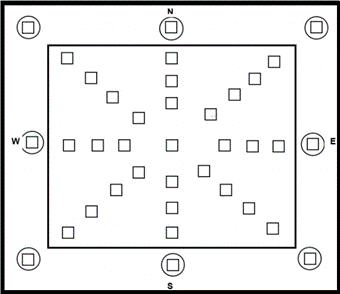
Los sitios de muestreo se ubicaron cada 75 m dentro del área de regeneración, siguiendo los puntos cardinales e intermedios norte, sur, este, oeste, noreste, sureste, suroeste y noroeste. Mediante la proyección de esas orientaciones hacia las áreas arboladas adyacentes, se ubicaron ocho sitios destinados a evaluar la diversidad de la vegetación arbórea y de la regeneración. La diversidad de la regeneración natural se evaluó en cuadrados de 5 x 5 m (25 m2), y la diversidad del arbolado mayor de 7.5 cm de diámetro, en círculos de 0.1 ha (Figura 2).
Análisis de datos
La información de campo, especies y número de individuos por especie se
utilizó para estimar los parámetros de diversidad alfa y beta. Los
estimadores aplicados para describir la diversidad alfa de las parcelas de
RAP, RAA y AA de cada una de las localidades fueron riqueza de especies
(S), definido como el número de especies presentes en
cada área de regeneración; índice de diversidad proporcional de Simpson (
Para estimar la diversidad beta se aplicó el índice cualitativo de Sørensen (Moreno, 2001):
donde,
A |
número de especies en el sitio A |
B |
número de especies en el sitio B |
C |
número de especies comunes a ambos sitios. |
Para determinar si existe diferencia significativa en la riqueza de especies entre las parcelas de RAP, RAA y AA por localidad se aplicaron análisis de rarefacción, mientras que, para la comparación de la diversidad de Shannon, diversidad de Simpson y equidad de Pielou, se aplicó la prueba estadística de t (α = 0.05) con el apoyo del programa Past 3.16 (Hammer, Harper, & Ryan, 2001).
Resultados
Especies
De las 30 especies reportadas por Medrano-Meraz et al. (2017) para la región de El Salto, P. cooperi, P. durangensis, P. strobiformis Engelm., P. leiophylla Schiede ex Schltdl. & Cham., P. teocote Schltdl. et Cham., P. herrerae Martínez, P. devoniana Lindley, Cupressus lindleyi Klotzsch ex Endl., Quercus rugosa Neé, Q. sideroxyla Bonpl. y Juniperus deppeana Steud. se encuentran presentes en el área de estudio, por lo que Alnus acuminata H. B. K. se adiciona a la lista de especies con presencia en esa región (Cuadro 1).
Cuadro 1 Especies presentes en las parcelas del área de estudio del ejido Pueblo Nuevo en la región de El Salto, Durango.
| Especies | RAP | RAA | AA |
|---|---|---|---|
| Pinus cooperi | X | X | X |
| Pinus durangensis | X | X | X |
| Pinus strobiformis | X | X | X |
| Pinus leiophylla | X | X | X |
| Pinus teocote | X | X | X |
| Pinus herrerae | X | X | X |
| Pinus devoniana | X | X | X |
| Cupressus lindleyi | - | - | X |
| Juniperus deppeana | X | X | X |
| Quercus rugosa | X | X | X |
| Quercus sideroxyla | X | X | X |
| Alnus acuminata | X | X | X |
RAP = Regeneración por árboles padres, RAA = Regeneración establecida bajo el dosel del arbolado adyacente, AA = Arbolado adulto adyacente al de la regeneración por árboles padres.
Descripción de los índices de diversidad
El número de especies presentes en las parcelas de RAP, RAA y AA fueron 11, 11 y 12, respectivamente. En el mismo orden de parcelas, los índices de diversidad proporcional de Shannon-Wiener fueron 1.53, 1.71 y 1.70; los de Simpson, 0.65, 0.72 y 0.73; y los de equidad de Pielou, 0.64, 0.71 y 0.69. A nivel de localidad, la riqueza de especies registrada fue 7, 4, 9, 9, 7 y 10 especies en Las Ciénegas, Ciénega Grande, El Venado, La Colmena, Los Bajíos y Pino Gordo, respectivamente. En el mismo orden de localidad, los indicadores de Shannon-Wiener fueron 1.18, 0.91, 1.31, 1.17, 1.52 y 1.67; el índice de Simpson tuvo valores de 0.55, 0.50, 0.62, 0.52, 0.75 y 0.78; y el de equidad de Pielou 0.60, 0.66, 0.59, 0.53, 0.78 y 0.73.
En el Cuadro 2 se presentan los indicadores de diversidad estimados en las parcelas de RAP, RAA y AA de cada una de las localidades. El número máximo de especies (S = 10) se registró en la parcela del AA de la localidad Pino Gordo, mientras que el número mínimo (S = 4), el cual fue igual en las tres parcelas evaluadas, correspondió a la localidad Ciénega Grande. Con relación al índice de Shannon, el valor mínimo (0.69) corresponde a las parcelas de RAP de la localidad Ciénega Grande y el mayor (1.68) a las del AA de la localidad Los Bajíos. A su vez, el índice de Simpson osciló entre 0.43 y 0.79; el mayor valor correspondió a la parcela de RAP de la localidad Pino Gordo. El índice de equidad de Pielou varió de 0.46 a 0.94; estos valores se registraron en las comunidades de El Venado y Los Bajíos, respectivamente, en las parcelas del AA.
Cuadro 2 Índices de diversidad de la regeneración en área de árboles padres (RAP), regeneración en áreas del arbolado adulto (RAA) y arbolado adulto (AA) adyacente por localidad en el ejido Pueblo Nuevo en la región de El Salto, Durango.
| Comunidad | Riqueza de especies | Índice de Shannon-Wiener | Índice de Simpson | Índice de Pielou | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RAP | RAA | AA | RAP | RAA | AA | RAP | RAA | AA | RAP | RAA | AA | |
| Las Ciénegas | 7 a | 6 a | 5 b | 1.32 a | 1.38 a | 0.78 b | 0.63 a | 0.71 a | 0.36 b | 0.68 a | 0.77 a | 0.49 b |
| Ciénega Grande | 4 a | 4 a | 4 a | 0.69 a | 1.20 b | 0.85 c | 0.44 a | 0.67 b | 0.47 c | 0.50 a | 0.86 b | 0.62 c |
| El Venado | 9 a | 8 a | 9 a | 1.46 a | 1.24 b | 1.00 c | 0.69 a | 0.63 b | 0.50 c | 0.67 a | 0.59 b | 0.46 c |
| La Colmena | 8 a | 7 a | 8 a | 0.99 a | 0.99 a | 1.23 b | 0.43 a | 0.45 a | 0.58 b | 0.48 a | 0.51 a | 0.59 b |
| Los Bajíos | 7 a | 5 b | 5 b | 1.59 a | 1.54 b | 1.68 c | 0.75 a | 0.70 b | 0.71 b | 0.76 a | 0.86 ab | 0.94 b |
| Pino Gordo | 6 a | 8 b | 10 c | 1.62 a | 1.46 b | 1.59 c | 0.79 a | 0.73 b | 0.75 c | 0.91 a | 0.70 b | 0.66 b |
Letras diferentes en cada índice de diversidad indican diferencia significativa (P < 0.05) entre las parcelas RAP, RAA y AA de cada comunidad.
Comparaciones de la diversidad alfa
La Figura 3 compara gráficamente la diversidad en las parcelas a nivel área de estudio. El análisis de rarefacción demostró que la riqueza de especies de los AA fue significativamente superior que la de RAP y RAA (P < 0.05). Las probabilidades de rechazo (P < 0.0001) estimadas con las pruebas estadísticas de t y la ausencia de traslapes entre los intervalos de confianza mostraron que la diversidad y la abundancia proporcional de especies de la RAP fueron significativamente inferiores a las que se presentan en las áreas de RAA y AA. A su vez, las diversidades proporcionales de Shannon-Wiener y Simpson de las parcelas RAA y AA fueron estadísticamente similares (P > 0.05) y mayores que en la RAP. Considerando el índice de Pielou, las parcelas de RAP fueron estadísticamente menos homogéneas que las de RAA y AA.

Figura 3 Estimadores puntuales e intervalos de confianza de los indicadores de diversidad. La diversidad en las parcelas de regeneración en árboles padres (RAP) es diferente de las de regeneración en arbolado adulto (RAA) y arbolado adulto (AA).
Los análisis de rarefacción realizados en cada una de las localidades demostraron que la riqueza de especies es similar entre las áreas RAP, RAA y AA en las localidades Ciénega Grande, El Venado y La Colmena, y que son diferentes en Las Ciénegas, Los Bajíos y Pino Gordo. En Las Ciénegas y Los Bajíos, la riqueza de especies fue estadísticamente superior (P < 0.05) que la de AA (Cuadro 2).
Por otra parte, de acuerdo con el Cuadro 3, los valores de t y la probabilidad de rechazo (P < 0.05) indican que las parcelas de RAP presentan indicadores de diversidad proporcional diferentes de las de RAA en las localidades Ciénega Grande, El Venado, Los Bajíos y Pino Gordo. De acuerdo con los índices de Shannon-Wiener y Simpson, la diversidad de las parcelas RAP y RAA contra AA son estadísticamente diferentes.
Cuadro 3 Estimadores de t y sus probabilidades de rechazo (P < 0.05) en la comparación de diferencias de los índices de diversidad de Shannon-Wiener y Simpson entre parcelas de cada localidad.
| Localidad | Parcelas | Shannon-Wiener | Simpson | ||
|---|---|---|---|---|---|
| Valor de t | Prob(t) | Valor de t | Prob(t) | ||
| Las Ciénegas | RAP vs RAA | -0.61 | 0.542 | 1.75 | 0.081 |
| RAP vs AA | 5.28 | 2.10 x 10-7 | -5.53 | 5.79 x 10-8 | |
| RAA vs AA | 4.06 | 6.08 x 10-5 | -2.94 | 0.003 | |
| Ciénega Grande | RAP vs RAA | -13.33 | 4.42 x 10-37 | 11.04 | 1.04 x 10-23 |
| RAP vs AA | -5.26 | 1.74 x 10-7 | 1.47 | 0.142 | |
| RAA vs AA | 10.94 | 1.1 x 10-25 | -12.43 | 5.65 x 10-33 | |
| El Venado | RAP vs RAA | 6.10 | 1.48 x 10-9 | -4.32 | 1.76 x 10-5 |
| RAP vs AA | 18.85 | 3.52 x 10-77 | -19.41 | 2.93 x 10-81 | |
| RAA vs AA | 6.10 | 1.40 x 10-9 | -8.92 | 1.39 x 10-18 | |
| La Colmena | RAP vs RAA | 0.07 | 0.942 | 0.94 | 0.346 |
| RAP vs AA | -9.18 | 5.25 x 10-20 | 12.32 | 1.65 x 10-34 | |
| RAA vs AA | -5.82 | 7.35 x 10-9 | 6.13 | 1.21 x 10-9 | |
| Los Bajíos | RAP vs RAA | 6.40 | 2.31 x 10-10 | -4.07 | 5.10 x 10-5 |
| RAP vs AA | 6.09 | 1.30 x 10-9 | -4.28 | 1.95 x 10-5 | |
| RAA vs AA | -2.33 | 0.020 | 0.80 | 0.425 | |
| Pino Gordo | RAP vs RAA | 5.54 | 4.22 x 10-8 | -6.69 | 4.77 x 10-11 |
| RAP vs AA | 3.26 | 0.001 | -11.58 | 1.12 x 10-30 | |
| RAA vs AA | -3.74 | 0.0002 | 1.99 | 0.047 | |
RAP = Regeneración en árboles padres, RAA = Regeneración en arbolado adulto, AA = Arbolado adulto.
En las Figuras 4 y 5 se observa que, de acuerdo con el índice de Shannon, la diversidad proporcional estimada en las parcelas de RAP, con respecto a la de RAA y AA, es superior en El Venado, Los Bajíos y Pino Gordo; es menor en Ciénega Grande; y es similar Las Ciénegas y La Colmena. Por su parte, el índice de equidad de Pielou demostró que el número de individuos por especies de la RAP es más homogéneo que el de las parcelas de RAA y AA en las localidades El Venado y Pino Gordo e igual de homogéneo en las comunidades Las Ciénegas, La Colmena y Los Bajíos.

Figura 4 Intervalos de confianza de los índices Shannon-Wiener y Pielou en las parcelas de las localidades Las Ciénegas, Ciénega Grande y el Venado. La ausencia de traslape indica diferencia significativa (P < 0.05) entre parcelas. RAP = regeneración en árboles padres, RAA = regeneración en arbolado adulto, AA = arbolado adulto.
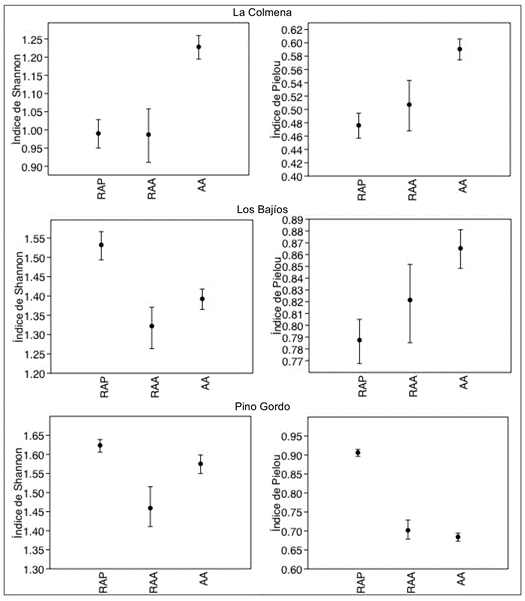
Figura 5 Intervalos de confianza de los índices Shannon-Wiener y Pielou en las parcelas de las localidades La Colmena, Los Bajíos y Pino Gordo. La ausencia de traslape indica diferencia significativa (P < 0.05) entre parcelas. RAP = regeneración en árboles padres, RAA = regeneración en arbolado adulto, AA = arbolado adulto.
Índice de similitud entre comunidades
El Cuadro 4 presenta el índice cualitativo de Sørensen que se estimó para valorar la similitud de especies entre las comunidades. Considerando las parcelas de RAP, el índice varió de 0.50 a 1.0. El valor más alto correspondió a Las Ciénegas y Los Bajíos, las cuales comparten el mismo número de especies; en contraste, el índice de similitud más bajo se obtuvo para Ciénega Grande y El Venado, quienes comparten solo tres especies. A su vez, los indicadores de similitud entre las comunidades en las parcelas de RAA osciló entre 0.50 a 0.91, donde el valor mayor correspondió a las localidades Las Ciénegas y Los Bajíos y el menor a El Venado y Los Bajíos. Los estimadores para valorar la similitud entre comunidades del AA variaron de 0.33 a 0.95; el valor menor pertenece a Ciénega Grande y El Venado, y el mayor se estimó en la dupla La Colmena y Pino Gordo.
Cuadro 4 Índices de similitud de la regeneración de árboles padres (RAP), regeneración en áreas adyacentes (RAA) y de la vegetación arbórea adyacente (AA) al área de árboles padres entre comunidades del ejido Pueblo Nuevo, El Salto, Durango.
| Comunidades contrastadas | Índices de similitud de Sørensen | ||
|---|---|---|---|
| RAP | RAA | AA | |
| Las Ciénegas vs Ciénega Grande | 0.73 | 0.80 | 0.89 |
| Las Ciénegas vs La Colmena | 0.88 | 0.71 | 0.71 |
| Las Ciénegas vs Venado | 0.67 | 0.77 | 0.62 |
| Las Ciénegas vs Los Bajíos | 1.00 | 0.91 | 0.80 |
| Las Ciénegas vs Pino Gordo | 0.92 | 0.71 | 0.67 |
| Ciénega Grande vs La Colmena | 0.62 | 0.67 | 0.62 |
| Ciénega Grande vs El Venado | 0.50 | 0.55 | 0.33 |
| Ciénega Grande vs Los Bajíos | 0.73 | 0.89 | 0.89 |
| Ciénega Grande vs Pino Gordo | 0.80 | 0.67 | 0.57 |
| El Venado vs La Colmena | 0.71 | 0.67 | 0.71 |
| La Colmena vs Los Bajíos | 0.88 | 0.77 | 0.71 |
| La Colmena vs Pino Gordo | 0.80 | 0.75 | 0.95 |
| El Venado vs Los Bajíos | 0.67 | 0.50 | 0.60 |
| El Venado vs Pino Gordo | 0.71 | 0.80 | 0.80 |
| Los Bajíos vs Pino Gordo | 0.92 | 0.77 | 0.67 |
Índices de similitud entre parcelas
Acorde con el Cuadro 5, los índices de similitud de Sørensen entre las parcelas variaron de 0.67 a 1.0, indicando que existe alta semejanza en especies entre las parcelas de RAP, RAA y AA. La cantidad de especies en las parcelas de RAP, con respecto a las de AA, fue mayor en Las Ciénegas y Los Bajíos; igual en Ciénega Grande, El Venado y La Colmena; e inferior en Pino Gordo. Estos resultados demuestran que la aplicación de la corta de árboles padres en bosques mixtos influye positivamente en la presencia de las especies nativas.
Cuadro 5 Índices de similitud de Sørensen entre parcelas por comunidad del ejido Pueblo Nuevo, El Salto, Durango.
| Localidad | Áreas | a | b | c | Sørensen |
|---|---|---|---|---|---|
| Las Ciénegas | RAP vs RAA | 7 | 6 | 6 | 0.92 |
| RAP vs AA | 7 | 5 | 5 | 0.83 | |
| RAA vs AA | 6 | 5 | 5 | 0.91 | |
| Ciénega Grande | RAP vs RAA | 4 | 4 | 4 | 1.00 |
| RAP vs AA | 4 | 4 | 4 | 1.00 | |
| RAA vs AA | 4 | 4 | 4 | 1.00 | |
| El Venado | RAP vs RAA | 9 | 8 | 7 | 0.94 |
| RAP vs AA | 9 | 9 | 9 | 1.00 | |
| RAA vs AA | 8 | 9 | 8 | 0.94 | |
| La Colmena | RAP vs RAA | 8 | 7 | 6 | 0.80 |
| RAP vs AA | 8 | 8 | 8 | 1.00 | |
| RAA vs AA | 7 | 8 | 6 | 0.80 | |
| Los Bajíos | RAP vs RAA | 7 | 5 | 5 | 0.83 |
| RAP vs AA | 7 | 5 | 5 | 0.83 | |
| RAA vs AA | 5 | 5 | 5 | 1.00 | |
| Pino Gordo | RAP vs RAA | 6 | 8 | 5 | 0.71 |
| RAP vs AA | 8 | 10 | 6 | 0.67 | |
| RAA vs AA | 8 | 10 | 8 | 0.89 |
a = número de especies de la comunidad A, b = número de especies de la comunidad B, c = número de especies comunes.
Discusión
Navar-Cháidez y González-Elizondo (2009) reportan que, en parcelas sujetas a tratamientos silvícolas, los indicadores de riqueza de especies y de diversidad de Shannon-Wiener y Simpson fueron iguales a 7, 0.76 y 0.73, respectivamente; estos valores son similares a los encontrados en las parcelas del presente estudio. Los indicadores a nivel de parcela son inferiores a los estimados a nivel regional, tal como lo señalan Medrano-Meraz at al. (2017), quienes indican que la riqueza de especies y la diversidad de Shannon-Wiener y Simpson fueron iguales a 30, 2.09 y 0.82, respectivamente.
La evaluación muestra que, en cinco de las seis localidades estudiadas, la riqueza de especies arbóreas en la RAP fue similar o mayor que en la RAA y AA (Cuadro 2). Leyva-López, Velázquez-Martínez, y Ángeles-Pérez (2010), en un estudio realizado en bosques mixtos en el estado de Oaxaca, reportaron resultados similares a los de este estudio. Estos autores argumentan que la presencia diferenciada de las especies puede estar relacionada con su habilidad de respuesta al nivel de disturbio. Por su parte, Smith et al. (1997) y Zavala (2001) mencionan que el número de especies está en función de la facultad que cada una de ellas tiene para reproducirse de manera vegetativa o regenerarse por semilla, así como de la vulnerabilidad al estrés causado por la incidencia directa de los rayos del sol, cambios de temperatura, disponibilidad de agua y competencia. Las prácticas de extracción del arbolado liberan al suelo de la cobertura de los árboles adultos y lo dejan expuesto a los rayos directos del sol, limitando el establecimiento de especies tolerantes a la sombra, pero favoreciendo la regeneración y desarrollo de las especies intolerantes (Smith et al., 1997). Por su parte, Sandor y Chazdon (2014) reportan que los árboles remanentes en las áreas manejadas explican la composición de especies de la regeneración establecida.
Con respecto al efecto que los métodos de regeneración han tenido en la riqueza de especies, Chaudhary, Burivalova, Koh, y Hellweg (2016) mencionan que los sistemas de selección y retención, donde se pueden incluir el sistema de protección y árboles padres, son calificados como los de menor impacto en el cambio de riqueza de especies, en contraste con el efecto que tiene el de matarrasa (Návar-Cháidez & González-Elizondo, 2009). Particularmente, el sistema de selección por árboles individuales permite el establecimiento y desarrollo solo de especies tolerantes a la sombra; el de selección en grupos a especies semitolerantes (Shields, Webster, & Nagel 2007); el de protección a especies semitolerantes e intolerantes a la sombra (Nasiri & Parsakhoo, 2012); y el de árboles padres a especies completamente intolerantes a la sombra (Smith et al., 1996). Considerando lo anterior, el registro de igual o mayor número de especies de la RAP en relación con el AA (Cuadro 2) se atribuye, en parte, a la presencia de especies de pino con capacidad de producir y dispersar semilla en los límites de las parcelas del AA y de las especies facultativas de reproducción vegetativa dentro de las parcelas de RAP. En este estudio, la presencia de las especies de los géneros Alnus y Quercus se atribuye a la facultad que tienen para reproducirse vegetativamente a partir de las raíces que quedan después del derribo, y la presencia de Pinus se debe a su capacidad de reproducirse por semilla; además, todas ellas responden favorablemente a la incidencia directa de los rayos del sol. Como ejemplos de la respuesta de los encinos a la reproducción vegetativa y de los pinos a la regeneración en áreas abiertas, Návar-Cháidez y González-Elizondo (2009) reportaron que P. cooperi, P. teocote, P. leiophylla, Juniperus spp. y Q. sideroxyla se establecieron exitosamente después de la aplicación de una corta de matarrasa aplicada en los bosques de Durango. Alanís-Rodríguez et al. (2012) también documentaron que las especies del género Quercus, en un ecosistema del noroeste de México, presentaron alta capacidad de rebrotar después de ser sujetas a disturbios causados por los incendios.
Los indicadores de Shannon y Simpson muestran que la diversidad proporcional de la RAP fue superior a la del AA en cuatro de las seis comunidades (Figuras 4 y 5), lo cual se atribuye al cambio en la abundancia proporcional de individuos por especie. Hernández-Salas et al. (2013) y Leyva et al. (2010) mencionan que la aplicación del método de árboles padres modifica la diversidad en la regeneración. Hernández-Salas et al. (2013) reportan que la aplicación periódica de las prácticas de manejo forestal, aunque aseguran la presencia de las especies, cambian la abundancia proporcional de las mismas, modificando los indicadores de diversidad. Al cambiar la abundancia proporcional de cada especie se modifica la composición y la diversidad proporcional (Del Río, Montes, Cañellas, & Montero, 2003). Pourmajidian et al. (2010) mencionan que la apertura del dosel, aparte de provocar cambios en la estructura, incrementa la diversidad.
En concordancia con los resultados de la diversidad alfa, el índice de diversidad beta indica que existe alta similitud de especies entre las parcelas de RAP, RAA y AA (Cuadro 5), demostrando que la selección de una sola especie como árboles padres en bosques mixtos no garantiza la dominancia y mucho menos la presencia única, como ocurre en los bosques caracterizados por una sola especie (Smith et al., 1997). Las especies que se encuentran en los límites y dentro del área de regeneración contribuyen a la riqueza de especies, bajo mecanismos de regeneración por semilla y reproducción por brotes. En estos casos, la cantidad de especies puede igualar e inclusive superar en número a las deseables, especialmente cuando en las áreas de regeneración se encuentran especies que se reproducen por brotes, pero que carecen de presencia en las áreas de arbolado adulto. En caso de desear controlar la composición, favoreciendo la presencia de una o varias especies, se debe plantear la aplicación de tratamientos complementarios como los aclareos, para manipular la proporción de especies deseables a través del control de la densidad (Louman, Quirós, & Nilsson, 2001). Leyva et al. (2010) y Hernández-Salas et al. (2013) afirman que el número de especies que se desarrollan dentro de una misma área disminuye al realizar tratamientos silvícolas. Contrario a lo anterior, cuando los árboles adyacentes sean intolerantes a la sombra, tengan edad para producir semilla y representen la diversidad de especies del bosque, la corta de árboles padres será una buena opción para aprovechar el recurso forestal maderable conservando la composición y diversidad del bosque.
Conclusiones
Los resultados sobre los análisis de riqueza y los índices de diversidad, equidad y similitud de especies demuestran que la corta de regeneración de árboles padres en bosques mixtos no afecta significativamente la composición y diversidad de las especies arbóreas presentes en la región, garantizando su conservación. El establecimiento de las especies y, por consiguiente, su preservación, se consiguen mediante la dispersión de la semilla producida por el arbolado adulto presente en los límites de las cortas de regeneración y por la reproducción vegetativa de las hojosas eliminadas dentro de las áreas de regeneración.
REFERENCIAS
Alanís-Rodríguez, E., Jiménez-Pérez, J., Valdecantos-Dema, A., González-Tagle, M. A., Aguirre-Calderón, O. A., & Treviño-Garza, E. T. (2012). Composición y diversidad de la regeneración natural en comunidades de Pinus-Quercus sometidas a una alta recurrencia de incendios en el noreste de México. Revista Mexicana de Biodiversidad, 83, 1208-1214. Retrieved from http://www.scielo.org.mx/pdf/rmbiodiv/v83n4/v83n4a26.pdf [ Links ]
Castellanos-Bolaños, J. F., Treviño-Garza, E. J., Aguirre-Calderón, O. A., Jiménez-Pérez, J., & Velázquez-Martínez, A. (2010). Diversidad arbórea y estructura espacial de bosques pino-encino en Ixtlán de Juárez, Oaxaca. Revista Mexicana de Ciencias Forestales, 1(2), 39-52. Retrieved from http://www.redalyc.org/articulo.oa?id=63438955004 [ Links ]
Chaudhary, A., Burivalova, Z., Koh, L. P., & Hellweg, S. (2016). Impacts of forest management on species richness: global meta-analysis and economic trade-off. Scientific Reports, 6(23954), 1-10. doi: 10.1038/srep23954 [ Links ]
Corral, R.J., Aguirre, C. O., Jiménez, P. J., & Corral, R. S. (2005). Un análisis del efecto del aprovechamiento forestal sobre la diversidad estructural en el bosque mesófilo de montaña El Cielo, Tamaulipas, México. Investigación Agraria: Sistema de Recursos Forestales, 14(2), 217-228. Retrieved from https://recyt.fecyt.es/index.php/IA/article/view/2278/1685 [ Links ]
Del Río, M., Montes, F., Cañellas, I., & Montero, G. (2003). Revisión: Índices de diversidad estructural en masas forestales. Investigación Agraria: Sistemas y Recursos Forestales, 12(1), 159-176. Retrieved from https://recyt.fecyt.es/index.php/IA/article/view/2490 [ Links ]
Gadow, K. V., Sánchez, S., & Aguirre-Calderón, O. (2004). Manejo forestal con bases científicas. Madera y Bosques, 10(2), 3-16. doi: 10.21829/myb.2004.1021271 [ Links ]
García, E. (1981). Modificaciones al sistema de clasificación climática de Köppen. México: Instituto de Geografía, Universidad Autónoma de México. [ Links ]
Grubb, P. J. (1977). The maintenance of species richness in plant communities. The importance of the regeneration niche. Biological Reviews, 52(1), 107-114. doi: 10.11117j.1469-185X.1977.tb01347.x [ Links ]
Hammer, Ø., Harper, D. A. T., & Ryan, P. D. (2001). PAST: Paleontological statistics software package for education. Retrieved March 9, 2018 from https://folk.universitetetioslo.no/ahammer/past [ Links ]
Hernández-Salas, J., Aguirre-Calderón, O., Alanís-Rodríguez, E., Jiménez-Pérez, J., Treviño-Garza, E., González-Tagle, M., & Domínguez-Pereda, L. (2013). Efecto del manejo forestal en la diversidad y composición arbórea de un bosque templado del noroeste de México. Revista Chapingo Serie Ciencias Forestales y del Ambiente, 19(2), 189-199. doi: 10.5154/r. rchscfa.2012.08.052 [ Links ]
Instituto Nacional de Estadística y Geografía (INEGI). (2010). Retrieved March 9, 2018 from https://www.inegi.org.mx/temas/mapas/edafologia/ [ Links ]
Ishii, H. T., Tanabe, S., & Hiura, T. (2004). Exploring the relationships among canopy structure, stand productivity and biodiversity of temperate forest ecosystems. Forest Science, 50(3), 342-355. Retrieved from http://www.researchgate.net/publication/233503484_Exploring_the_Relationships_Among_Canopy_Structure_Stand_Productivity_and_Biodiversity_of_Temperate_Forest_Ecosystems [ Links ]
Jayakumar, R., & Nair, K. K. N. (2013). Species diversity and regeneration patterns in tropical forests of the Western Ghats, India. ISRN Ecology, Article ID 890862, 1-14. doi: 10.1155/2013/890862 [ Links ]
Khumbongmayum, A. D., Khan, M. L., & Tripathi, R. S. (2005). Survival and growth of seedlings of a few species in four sacred groves of Manipur, Northeast India. Current Science, 88(11), 1781-1788. Retrieved from https://www.researchgate.net/publication/236628069_Survival_and_growth_of_seedlings_of_a_few_tree_species_in_the_four_sacred_groves_of_Manipur_Northeast_India [ Links ]
Leyva-López, J. C., Velázquez-Martínez, A., & Ángeles-Pérez, G. (2010). Patrones de diversidad de la regeneración natural en rodales mezclados de pinos. Revista Chapingo Serie Ciencias Forestales y del Ambiente, 16(2), 227-239. doi: 10.5154/r.rchscfa.2010.06.038 [ Links ]
Louman, B., Quirós, D., & Nilsson, M. (2001). Silvicultura de bosques latifoliados húmedos con énfasis en América Central. Turrialba, Costa Rica: CATIE. [ Links ]
Malik, Z. A., & Bhatt, A. B. (2016). Regeneration status of tree species and survival of their seedlings in Kedarnath Wildlife Sanctuary and its adjoining areas in Western Himalaya, India. Tropical Ecology, 57(4), 677-690. Retrieved from https://www.researchgate.net/publication/313368616_Regeneration_status_of_tree_species_and_survival_of_their_seedlings_in_kedarnath_wildlife_sanctuary_and_its_adjoining_areas_in_Western_Himalaya_India [ Links ]
Medrano-Meraz, M. J., Hernández, F. J., Corral-Rivas, S., & Nájera-Luna, J. A. (2017). Diversidad arbórea a través de gradientes topográficos en la Región de El Salto, Durango. Ecosistemas y Recursos Agropecuarios, 4(12), 525-534. doi: 10.19136/era.a4n12.1121 [ Links ]
Moreno, E. C. (2001). Métodos para medir la biodiversidad (vol. 1). Zaragoza, España: M&T-Manuales y Tesis SEA. [ Links ]
Nasiri, M., & Parsakhoo, A. (2012). Shelterwood cutting system for forest management. Journal of Applied Biological Sciences, 6(3), 57-60. Retrieved from http://dergipark.gov.tr/download/article-file/415652 [ Links ]
Návar-Cháidez, J. J., & González-Elizondo, S. (2009). Diversidad, estructura y productividad de bosques templados de Durango, México. Polibótanica, 27, 71-87. Retrieved from http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S1405-27682009000100005 [ Links ]
Pourmajidian, M. R., Jaliluand, H., Fallah, A., Husseini, S. A., Parskhoo, A., Vosoughian, A., & Rahmani, A. (2010). Effect of shelterwood cutting method on forest regeneration and stand structure in a Hyrcanian forest ecosystem. Journal of Forestry Research, 2(3), 265-272. doi: 10.1007/s11676-010-0070-7 [ Links ]
Sandor, M. E., & Chazdon, R. L. (2014). Remnant trees affect species composition but not structure of tropical second-growth forest. PloS ONE, 9(1), e83284. doi: 10.1371/journal.pone.0083284 [ Links ]
Shields, J. M., Webster, Ch. R., & Nagel, L. M. (2007). Factors influencing tree species diversity and Betula alleghaniensis establishment in silvicultural openings. Forestry, 80(3), 293-307. doi: 10.109/forestry/cpm013 [ Links ]
Smith, M. D., Larson, B. C., Kelty, M. J., & Ashton, P. M. S. (1997). The practice of silviculture applied forest ecology (9th ed.). USA: John Wiley & Sons. [ Links ]
Solís, M. R., Aguirre, C. O., Treviño, G. E., Jiménez, P. J., Jurado, Y. E., & Corral-Rivas, J. (2006). Efecto de dos tratamientos silvícolas en la estructura de ecosistemas forestales en Durango, México. Madera y Bosque, 12(2), 49-64. Retrieved from http://www.redalyc.org/articulo.oa?id=61712205 [ Links ]
Zavala, Ch. F. (2001). Introducción a la ecología de la regeneración natural de encinos. México: Universidad Autónoma Chapingo. [ Links ]
Agradecimientos
Se agradece al Tecnológico Nacional de México por el apoyo otorgado a través del Instituto Tecnológico de El Salto, así como a la Unidad de Servicios Técnicos del ejido de Pueblo Nuevo por las facilidades prestadas para la realización de este proyecto.
Recibido: 02 de Septiembre de 2018; Aprobado: 09 de Marzo de 2019
 This is an open-access article distributed under the terms of the
Creative Commons Attribution License
This is an open-access article distributed under the terms of the
Creative Commons Attribution License
