











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
The Coyote (Canis latrans) is a predator that is considered to have a high degree of ecological plasticity; consequently, it can be found in different types of ecosystems including suburban and urban human settlements. Its geographical distribution ranges from North America to northern Panamá (Bekoff and Gese 2003; Marín et al. 2015; Méndez-Carvajal and Moreno 2014; Hody et al. 2019). The coyote's reproductive success and its ability to disperse, has allowed it to have high population numbers and a wide distribution (Carreón 1998; Garrido and Arribas 2008; Hernández and Laundré 2014; Méndez-Carvajal and Moreno 2014).
The home range of the coyote may comprise 3 km2 to 67 km2 and its extension varies according to the characteristics of the individual and the stage of life in which it is found (Hernández 1990; Hernández et al. 1993; Servín and Huxley 1995; Hidalgo-Mihart et al. 2001). It has been hypothesized that its wide distribution and population abundance in North America is due to the elimination of the Gray wolf (Canis lupus), since both canids were sympatric species competing for resources. Additionally, it has been documented that C. lupus preyed on C. latrans (Hall 1981; Vaughan and Rodríguez 1986).
Although the coyote is a carnivore, the modification of its diet is an adaptation to the different environments in which it is found, adopting an omnivorous diet. All these characteristics together, have led it to be a successful predator (Bekoff 1977; Hall 1981; Vaughan and Rodríguez 1986). The coyote is considered an important regulator of populations of mammals and insects (Andelt et al. 1987), rodents and lagomorphs. Additionally, they consumed seeds from their herbivorous diet, fulfilling a relevant role in the regeneration of pine-oak forests. Therefore, the eradication of coyote populations would have a drastic effect on animal and plant communities and on ecosystems as a whole (Servín and Huxley 1993, 1995).
The coyote is considered an abundant and widely distributed species in Mexico, however there are only a few studies on its relative abundance, distribution, role they play in a given habitat, or diet in any ecosystem in Chihuahua, northern Mexico (Pérez et al. 1982; Vela 1985; Pacheco et al. 1999). Therefore, our objective was to estimate and compare relative abundance index, habitat selection, and diet of coyote (C. latrans) in two types of vegetation (semi-open grassland and pine-oak forest) in northern Mexico, through indirect methods such as scent stations and scats. The information generated might be use by local authorities and Rancho Experimental Teseachi managers to carry out conservation plans.
Materials and methods
The study was conducted at Rancho Experimental Teseachi, located in central-western Chihuahua. The ranch is owned by Universidad Autónoma de Chihuahua, and has as its main objective teaching, research, and transfer of technology in animal science and natural resources (Espinoza and Quintana 2013; Álvarez-Córdova et al. 2019). It encompasses approximately 12,300 ha and is located between the municipalities of Namiquipa, Bachíniva, and Guerrero (28° 53’ 44” N, -107° 27’ 22” W) at 2,250 masl (Figure 1). The landscape is shaped by hills and high mountain ranges displaying a mixture of arboreal pasture grasses, oak chaparral, temperate forest, semi-open grasslands, and pine-oak forest (COTECOCA 1978; Álvarez-Córdova et al. 2019). For this study, we selected the vegetation pine-oak forest and semi-open grassland, due to the previous information of the ranch staff and personal observations of the presence of coyotes in both areas.
From April 2018 to March 2019, we placed 10 fixed scent stations (SS) in pine-oak forest and 10 SS in semi-open grassland. A five-kilometer transect with a separation of 500 linear meters (Figure 1) between each scent stations was established in each type of vegetation. The SS consisting of 1.00 meter in diameter, where vegetation was removed and sand or soil fine were added and were baited with chicken and sardine (Linhart and Knowlton 1975). The tracks of species visited the stations at night were recorded in the SS. All scent stations were activated during the afternoon and checked the next day in the morning (two nights / month; Linhart and Knowlton 1975; Roughton and Sweeny 1982; Conner et al. 1983; Carreón 1998).
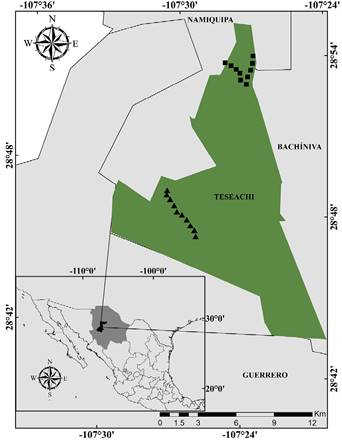
Figure 1 Geographic location of Rancho Experimental Teseachi, Chihuahua, Mexico. Black squares and black triangles indicate sampling sites in semi-open grassland and pine-oak forest respectively.
Traces and scats we found at SS and through the linear transect were identified as belonging to C. latrans following Aranda (2012). Each collected scat was georeferenced through a Global Positioning System (GPS; Garmin GPSMAP 66s), measurements (width and length) were taken with a measuring tape, and they were subsequently stored individually in airtight bags (Álvarez-Córdova et al. 2019). Once in the laboratory they were cleaned according to Arnaud (1993) and Álvarez-Córdova et al. (2019). Following the field guidelines of Álvarez-Castañeda et al. (2015), Aguirre-Segura and Barranco (2015) and Lebgue et al. (2015) the items within the scats were identified.
To estimate the relative abundance index, we generated a database with the records of visits of C. latrans at the SS, where coyote records are organized for each sampling. We defined a visit as the presence of at least one trace of coyote. To calculate the relative abundance index (RAI) we used the formula proposed by Linhart and Knowlton (1975): number of visits of the study species / number active of scent stations per night (Linhart and Knowlton 1975; Roughton and Sweeny 1982; Conner et al. 1983; Diefenbach et al. 1984; Carreón 1998). The records were analyzed on an annual and seasonal basis.
We performed a chi/square test to compare patterns of habitat selection between pine-oak forest and semi-open grassland. To achieve this, the information collected was divided between records of C. latrans presence and absence in scent stations in both types of vegetation. It was taken as a null hypothesis (H0) that relative abundance index of C. latrans was independent on the type of vegetation (α = 0.05). A double entry contingency table was developed, in which the observed frequencies (OF) corresponded the records of scent stations that were positive during the study in the two types of vegetation and expected frequencies (EF) for forest and grassland were calculated according to chi-square formula. The results were analyzed annually and seasonally between both types of vegetation. To describe the diet of coyote, we separated the food items, and a percentage of frequency was calculated for each item (Linhart and Knowlton 1975; Roughton and Sweeny 1982; Conner et al. 1983; Carreón 1998; Monroy-Vilchis and Velázquez 2002).
Results
We obtained 113 coyote visits in total of 420 scent stations placed during an annual period, 67 visits correspond to pine-oak forest and 46 to semi-open grassland. The annual RAI in pine-oak forest was 0.30 versus semi-open grassland with 0.23 (Figure 2). We found differences in the RAI among seasons in both ecosystems, indicating more RAI of coyotes during Spring (0.37) and Autumn (0.40) compared to summer and winter. In semi-open grassland the highest RAI was found in Autumn (0.30). There were differences between seasons, nevertheless, regardless of the season, the RAI was always greater in pine-oak forest than in semi-open grassland (Figure 2).
For habitat selection, the double entry contingency table showed that of 420 scent stations (200 SS for grassland and 220 SS for forest) set during sampling, in 113 SS the presence of C. latrans were registered, corresponding to frequencies observed of 46/113 and 67/113 for semi-open grassland and pine-oak forest, respectively (Table 1). However, we did not find a significant difference between both ecosystems during the year (χ 2 = 2.96, P > 0.05). In the same way, we did not find significant differences (all P > 0.05) between seasons, Spring (χ 2 = 3.26), Summer (χ 2 = 2.63), Autumn (χ 2 = 0.87) and Winter (χ 2 = 0.89). Therefore, the results suggest that the relative abundance index of coyotes and type of vegetation are independent variables across seasons.
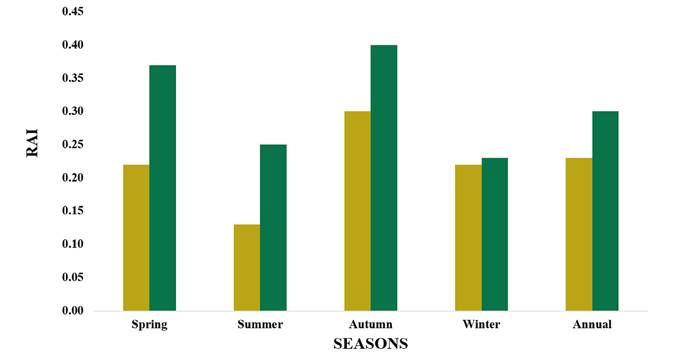
Figure 2 Relative abundance index (RAI) of Canis latrans seasonally and annually during 2018-2019 at Rancho Experimental Teseachi, Chihuahua, Mexico. Semi-open grassland (gray bars) and pine-oak forest (black bars).
We collected a total of 34 scats, four in semi-open grassland and 30 for pine-oak forest (2.0 to 3.1 [2.4] ± 0.33 cm width x 13.0 to 15.0 [13.3] ± 0.87 cm length). Annually, 55.8 % the food items recovered from the scats belong to mammals, 35.3 % to plants and 8.8 % to insects (Orthoptera: Acrididae). For Spring season, 16 scats were collected, in which 68.8% of the food items recovered belonged to mammals and 31.2 % to plants. During Summer, seven scats were collected, with 71.4 % of the remains corresponding to mammals and 28.6 % to insects. In Autumn, only five scats were recovered where 40.0 % of the items belonged to mammals, 40.0 % to plants and 20.0 % to insects. Finally, in Winter six scats were found and 16.7 % of the items recovered were mammals and 83.3 % plants. The mammalian species identified corresponded to Cotton rats (Sigmodon sp.), Woodrats (Neotoma sp.), and Cottontail rabbits (Sylvilagus sp). The plants recovered corresponded to fruits of Manzanita (genus Arctostaphylos), and the insects were grasshoppers of the family Acrididae (Table 2). The four scats collected in semi-open grassland corresponded only to plants.
Table 1 Double entry contingency table (χ 2; OF: observed frequencies, EF: expected frequencies) for Canis latrans at Rancho Experimental Teseachi, Chihuahua, Mexico.
| Vegetation | Presence | Absence | Total |
|---|---|---|---|
| Semi-open grassland | OF = 46.0 EF = 53.8 | OF = 154.0 EF = 146.2 | 200 |
| Pine-oak forest | OF = 67.0 EF = 59.2 | OF = 153.0 EF = 160.8 | 220 |
| Total | 113 | 307 | 420 |
Discussion
Throughout the sampling period, the pine-oak forest showed the greatest relative abundance index of C. latrans. This may be because the forest provides them with shelter from climatic situations and a greater variety of seeds, fruits, and insects. Differences in the RAI between pine-oak forest and semi-open grassland were found and this result is in agreement with Monroy-Vilchis and Velázquez (2002), who found the highest RAI of C. latrans in pine and mixed forests. However, our results differ from those of Ponce et al. (2005), who found coyote lower RAI in forests, maybe because of the complexity of capturing prey, and competition with other predators (mountain lions Puma concolor and black bears Ursus americanus).
Table 2 Annual and seasonal variation in diet of Canis latrans at Rancho Experimental Teseachi, Chihuahua, Mexico.
| Season | Mammals | Plants | Orthoptera | Scats |
|---|---|---|---|---|
| Spring | 68.7 % | 31.2 % | 0 % | 16 |
| Summer | 71.4 % | 0 % | 28.6 % | 7 |
| Autumn | 40.0 % | 40.0 % | 20.0 % | 5 |
| Winter | 16.7 % | 83.3 % | 0 % | 6 |
| Annual | 55.9 % | 35.3 % | 8.8 % | 34 |
Several authors have recognized forest and grassland as important ecosystems for mammals that use them for protection and food (Servín and Huxley 1991; Hidalgo-Mihart et al. 2001; Randa and Yunger 2004). In this study the two seasons with the highest RAI were Spring and Autumn in both ecosystems, which are the last seasons coinciding with an increase in rainfall, resulting in a higher abundance of potential prey (rodents and lagomorphs), and other food items like fruits. According to the yearly records, Autumn seems to be a key season for coyote populations because of the dispersal of young (Randa and Yunger 2004).
Although the RAI was different between pine-oak forest and semi-open grassland, the habitat selection test did not show statistically significant differences in annual relative abundance index of coyotes between the two vegetation types. This could affirm that the coyote did not select a particular type of vegetation. Furthermore, the season that presented the greatest variation was Spring, while Autumn showed less variation. Although different authors reported that potential prey and vegetation defines C. latrans presence in different ecosystems, we believe that food is a key variable for the presence of coyotes in any ecosystem (Bekoff 1977; Hall 1981; Vaughan and Rodríguez 1983; Bekoff and Gese 2003; Randa and Yunger 2004; Ponce et al. 2005).
In the annual diet, mammals were the main food source with 55.8 %, followed by plants (35.3 %). We only know of two studies related to the coyote diet in Chihuahua. Pérez et al. (1982) and Vela (1985) reported that mammals (rodents and lagomorphs) are the main food item of the coyote diet, while vegetable matter was consumed occasionally in central Chihuahua. Other studies carried out in Durango reported similar food preferences, adding insects as an important food item during the dry season (Servín and Huxley 1991; Grajales-Tam and González-Romero 2014). Diet preferences coincide with the ecosystem role this canid plays as a biological control of rodent and lagomorphs populations, and seed disperser in different ecosystems in which it is distributed (Servín and Huxley 1991; Grajales-Tam et al. 2003; Cruz-Espinoza et al. 2008; Arias-Del Razo et al. 2011; Grajales-Tam and González-Romero 2014; Poessel et al. 2017).
We found differences in diet between seasons, which can be explain by the variation in food availability among seasons (Randa and Yunger 2004; Ponce et al. 2005). The small rodents and lagomorphs consumed by C. latrans can be found throughout the four seasons (Servín and Huxley 1991); nevertheless, fruits are found in Spring, Autumn and Winter; and Orthopteran insects only appear in Summer and Autumn, corresponding to the rainy season when there is an increase in the population of Acrididae (Rivera 2006). Coyotes need the greatest energy intake for reproduction in the winter months, so that pups will be born in spring coinciding with the yearly food peak (Roughton and Sweeny 1982; Servín and Huxley 1991; Bekoff and Gese 2003; Randa and Yunger 2004; Ponce et al. 2005; Hernández and Laundré 2014). Even though scent stations are a very low-cost sampling technique, it has been overshadowed by other techniques such as camera traps and radiotelemetry due to the quantitative information they provide. However, the scent stations showed an efficacy in the evaluation of the relative abundance index of coyote in this study and with the data it was possible to make comparisons between ecosystems, being functional to determine population fluctuations.
In addition, in the collected scats there were specimens of a genus of nematode (Physaloptera sp.) not reported before for the state of Chihuahua, México (Álvarez-Córdova et al. 2019). The presence of this nematode in scats of the coyote agrees with the feeding habits of this canid, because this nematode is parasite of intermediate hosts like lagomorphs and rodents (Álvarez-Córdova et al. 2019). This study was conducted systematically in order to generate information about the ecology of Canis latrans in Chihuahua, specifically about the relative abundance index of this carnivore in two types of vegetation in an annual period for central-western Chihuahua populations. In addition, it is corroborated that coyote rather than a selective carnivore is an opportunistic carnivore in its diet, so it fulfills its function as a biological control of different species.

