











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Neomillspaughia y Podopterus son dos géneros de hábito leñoso cuya distribución se restringe a las selvas caducifolias de México y América Central (Brandbyge, 1993; Burke et al., 2010; Hernández-Ledesma et al., 2015). Cada uno de los géneros tiene tres especies descritas que poseen en común tépalos acrescentes en el fruto con alas dorsales. Neomillspaughia se diferencia por tener flores en panículas y tépalos exteriores con las alas dorsales sin extenderse a los pedicelos, mientras que en Podopterus las flores son fasciculadas y los tépalos exteriores con las alas dorsales decurrentes en el pedicelo (Blake, 1921). El número de tépalos es una característica clave para el reconocimiento de ambos géneros; por un lado, cinco tépalos es un número consistente en Neomillspauhia (Blake, 1921; Standley y Steyermark, 1946) mientras que en Podopterus suele variar de cinco a seis (Blake, 1921; Burger, 1983; Ortiz-Díaz, 1994; Standley y Steyermark, 1946) pudiendo ocasionalmente, presentarse cuatro. Las relaciones filogenéticas de estos dos géneros son controversiales resolviéndose Neomillspaughia como grupo hermano de Coccoloba ambos con cinco tépalos y Podopterus en una posición más afín a los géneros con seis tépalos como Gymnopodium, Leptogonum, Ruprechtia y Triplaris (Burke et al., 2010). No obstante, Burke y Sanchez (2011) incluyen a Podopterus dentro de la tribu Coccolobeae junto con Coccoloba y Neomillspaughia. Aunque las diferencias morfológicas entre Neomillspaughia y Podopterus son sustanciales, el número de especies empleadas en estudios filogenéticos moleculares se limita a tres, N. emarginata, P. mexicanus y P. cordifolius de tal manera que las relaciones filogenéticas en la subfamilia Eriogonoideae aún son inciertas.
Estudios palinológicos en Polygonaceae realizados por diversos autores (Brandbyge, 1990b, 1993; Brandbyge y Øllgaard, 1984; Nowicke y Skvrla, 1977) en la subfamilia Eriogonoideae mencionan que el polen presenta poca variación, sin embargo, existe una organización diversa de los aperturas y ornamentaciones entre los géneros de esta subfamilia (Burke et al., 2010; Mondal, 1997) lo que les confiere interés taxonómico. El polen en la subfamilia es tri, o menos comúnmente, tetracolporado, la exina varía de punctada y punctado-estriada a reticulada y microreticulada, así como también espinulosa.
Aunque la utilidad taxonómica del polen para explicar las relaciones filogenéticas en Eriogonoideae ha sido limitada (Burke et al., 2010), hay que señalar que su potencial no ha sido explorado en todas las especies de los géneros Neomillspaughia y Podopterus. Por ejemplo N. hondurensis descrita por Ortiz-Díaz et al. (2013) no se ha considerado en estudios filogenéticos ni palinológicos previos. La variación en la forma, número de aperturas, así como también en la ornamentación de la exina de los granos de polen podrían sugerir relaciones con otros géneros de Eriogonoideae. Los objetivos de este estudio son caracterizar el polen de Neomillspaughia y Podopterus y explorar el valor taxonómico del polen en la delimitación de sus especies y sus afinidades con otros géneros de la subfamilia Eriogonoideae.
Materiales y métodos
Selección de las especies
Para el estudio palinológico se consideraron todas las especies de Neomillspaughia y Podopterus: N. emarginata, N. hondurensis, N. paniculata, P. cordifolius, P. guatemalensis y P. mexicanus. En total se seleccionaron 16 ejemplares de herbario (Apéndice) procedentes de los herbarios del Museo de Historia Natural de Londres (BM), Universidad Nacional Autónoma de México (MEXU), Jardín Botánico de Missouri (MO) y de la Universidad Autónoma de Yucatán (UADY). Para las especies N. emarginata y N. paniculata se analizaron 4 ejemplares de cada uno y de N. hondurensis, tres pliegos de herbario. Para las especies de P. mexicanus y P. cordifolius dos, y para P. guatemalensis, uno.
Técnica de acetólisis
El polen se acetolizó de acuerdo con la técnica de acetólisis (Erdtman, 1952) siguiendo las adecuaciones de Alfaro-Bates et al. (2010). Los granos de polen obtenidos se colocaron en gelatina glicerinada (Sawyer, 1988) haciendo las preparaciones permanentes, mismas que después de su descripción se depositaron en la colección palinológica de la Universidad Autónoma de Yucatán (UADY-PAL).
Medición y descripción de los granos de polen
Para el registro de las mediciones se tomaron en cuenta 30 granos de polen de cada especie de Neomillspaughia y 14 granos de polen para cada especie de Podopterus. Esto se justifica debido a la disponibilidad de ejemplares de herbario y a que algunos ejemplares presentaron pocos granos de polen. Aun así, el esfuerzo de observación y medición cumple con el número estándar en las descripciones palinológicas de 10-15 granos por especie como en las presentadas por Hong et al. (2005), Yasmin et al. (2009, 2015) o Yurtseva et al. (2014). Las observaciones, mediciones y fotografías se tomaron en las vistas polar y ecuatorial empleando un microscopio óptico Zeiss Primo Star y cámara Infinity3 a 1000 aumentos. Las mediciones se tomaron en las microfotografías digitales previa calibración en el programa ImageJ ver. 1.44p (Schneider et al., 2012). El promedio y desviación estándar de los datos se expresan en µm. En vista ecuatorial, la medición del eje polar (P) y el eje ecuatorial (E) permite obtener la relación (P/E) y así determinar la forma del grano de polen en vista ecuatorial. En vista polar, se midió la distancia entre los colpos (dc) y el diámetro (d) y con estos valores se obtiene el índice de área polar (dc/d). Para la observación detallada de la ornamentación de la exina se empleó el microscopio electrónico de barrido (MEB) JEOL JSM-6360LV. Para ello, los granos acetolizados fueron deshidratados en un tren de alcohol de 30%, 50% y 70%, montados sobre cinta de carbón bifaz y recubiertos con oro. El microscopio electrónico de barrido fue operado a 20kV. La terminología utilizada para la descripción palinológica de las especies incluye la de Erdtman (1952), Sáenz Laín (2004) y Punt et al. (2007).
Resultados
Neomillspaughia S. F. Blake, Bull. Torrey Bot. Club 48(3): 84-85. 1921.
Granos radiosimétricos isopolares, prolato a subprolatos dada la relación eje polar/ecuatorial de 1.32 a 1.36; tricolporados. Tectum perforado y microrreticulado, con exina de 0.64-1.07 µm de grosor. Endoaperturas lalongadas; colpos con margen o vestíbulo. Índice de área polar pequeña.
Neomillspaughia emarginata (H. Gross) S. F. Blake, Bull. Torrey Bot. Club 48(3): 85. 1921 (Láminas I, II y III A).
Granos tricolporados, prolatos de 12.97±0.85 µm × 9.76±1.49 µm. Relación P/E= 1.36. Exina de 0.90±0.17 µm de grosor, sexina de 0.53 µm de espesor y nexina de 0.37 µm. Tectum microrreticulado. Colpos de 4.30±0.80 × 1.27±0.34 µm con margen. Endoaperturas lalongadas de 4.85±0.98 µm de largo × 1.32±0.43 de µm de ancho. Granos de tamaño pequeño según el índice de área polar= 0.24.
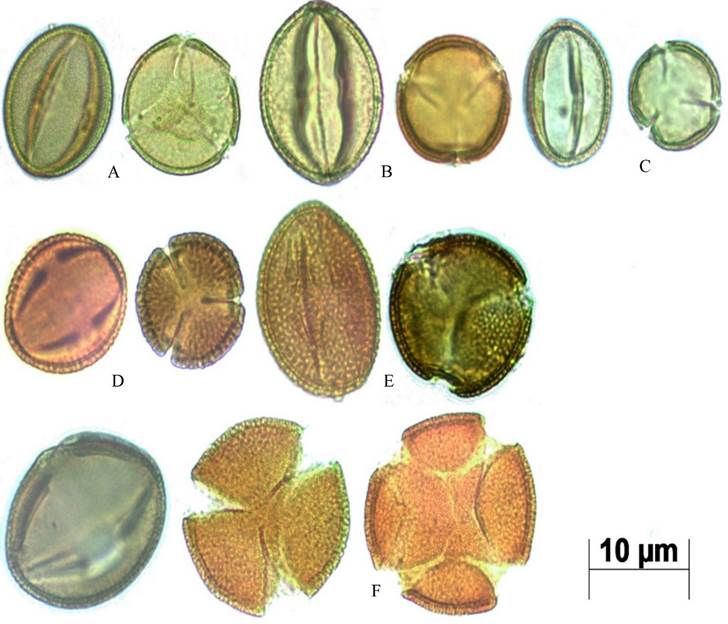
Lámina I: Microfotografías de granos de polen de Neomillspaughia y Podopterus en vista ecuatorial y polar tomadas con microscopio óptico a 1000 aumentos. A. Neomillspaughia emarginata; B. N. hondurensis; C. N. paniculata; D. Podopterus cordifolius; E. P. guatemalensis; F P. mexicanus con granos tri-tetracolporados.

Lámina II Vistas polar y ecuatorial de los granos de polen de Neomillspaughia y Podopterus tomadas con el MEB. A. Neomillspaughia emarginata en vista subecuatorial; B. N. hondurensis (ecuatorial); C. N. paniculata (subecuatorial); D. Podopterus cordifolius vista polar; F. P. guatemalensis (subpolar); G. P. mexicanus (polar).
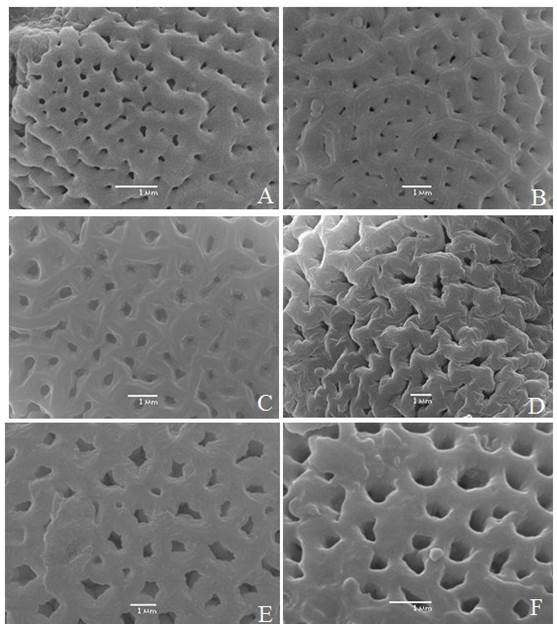
Lámina III Microfotografías de la superficie de la ectexina de Neomillspaughia y Podopterus tomadas con el MEB. A. Neomillspaughia emarginata B. N. hondurensis; C. N. paniculata; D. Podopterus cordifolius; E. P. guatemalensis; F. P. mexicanus.
Neomillspaughia hondurensis J.J. Ortíz & Arnelas, Phytotaxa 144(2): 57-60, f. 1C-D. 2013. (Láminas I, II y III B)
Granos tricolporados, subprolatos de 15.34±1.45 µm × 11.76±1.64 µm. Relación P/E= 1.32. Exina de 0.90±0.15 µm de grosor, sexina de 0.51 µm de espesor y nexina de 0.39 µm. Tectum microrreticulado. Colpos de 5.62±1.92 µm × 1.20±0.27 µm con vestíbulo. Endoaperturas lalongadas de 7.60±2.47 µm de largo x 1.59±0.34 µm de ancho. Granos de tamaño pequeño según el índice de área polar= 0.19.
Neomillspaughia paniculata (Donn. Sm.) S. F. Blake, Bull. Torrey Bot. Club 48(3): 85. 1921. (Láminas I, II y III: C).
Granos tricolporados, prolatos de 9.81±2.87 µm × 7.41±2.25 µm. Relación P/E= 1.34. Exina de 0.84±0.20 µm de grosor, sexina de 0.48 µm de espesor y nexina de 0.36 µm. Tectum microrreticulado. Colpos de 3.45±0.66 µm × 1.12±0.21 µm con margen. Endoaperturas lalongadas de 5.26±1.57 µm de largo × 1.25±0.24 µm de ancho. Granos de tamaño pequeño según el índice de área polar= 0.21.
Podopterus Pl. Aequinoct. 2: 89. 1809[1812].
Granos radiosimétricos isopolares, subprolatos a prolato esferoidales dada la relación eje polar/ecuatorial de 1.11 a 1.30; tricolporados (rara vez, tetracolporados y en proporción variable). Tectum perforado y microrreticulado en su mayoría, con exina de 0.95-1.41 µm de grosor. Endoaperturas lalongadas o circulares; colpos con margen o vestíbulo. Índice de área polar pequeña a mediana.
Podopterus cordifolius Rose & Standl. Proc. Biol. Soc. Washington 33: 66. 1920. (Láminas I, II y III D)
Granos tricolporados, prolato esferoidales de 12.34±1.11 µm × 11.15±1.16 µm. Relación P/E= 1.11. Exina de 1.19±0.22 µm de grosor, sexina de 0.73 µm de espesor y nexina de 0.46 µm. Tectum reticulado con lúmenes más profundos y definido que en el resto de las especies. Colpos de 4.79±0.92 µm × 1.52±0.44 µm con margen. Endoaperturas lalongadas de 6.61±0.93 µm de largo × 1.18±0.43 µm de ancho. Granos de tamaño pequeño según el índice de área polar= 0.22.
Podopterus guatemalensis S. F. Blake, Bull. Torrey Bot. Club 48(3): 87. 1921. (Láminas I, II y III E).
Granos tricolporados, subprolatos de 15.97±1.29 µm × 12.30±0.58 µm. Relación P/E= 1.30. Exina de 1.08±0.01 µm de grosor, sexina de 0.59 µm de espesor y nexina de 0.50 µm. Tectum micorreticulado. Colpos de 6.12±1.43 µm × 2.37±0.51 µm con vestíbulo. Endoaperturas lalongadas de 7.54±1.83 µm de largo × 1.69±0.19 µm de ancho. Granos de tamaño pequeño según el índice de área polar= 0.24.
Podopterus mexicanus Bonpl. Pl. Aequinoct. 2: 89-91, t. 107. 1809[1812]. (Láminas I, II y III F).
Granos tricolporados o tetracolporados, subprolatos de 17.13±1.23 µm × 13.82±1.62 µm.
Relación P/E=1.25. Exina de 1.12±0.17 µm de grosor, sexina de 0.64 µm de espesor y nexina de 0.48 µm. Tectum microrreticulado. Colpos de 5.55±0.64 µm × 2.49±0.94 µm con margen. Endoaperturas circulares de 5.61±0.81 µm de diámetro. Granos de tamaño mediano según el índice de área polar= 0.27.
Cuadro 1 Cuadro resumen con los caracteres palinológicos evaluados en las especies estudiadas. EP= Eje Polar; EE.= Eje Ecuatorial; RP/E= Relación Ejes Polar y Ecuatorial; Forma, I= Subprolato, II= Prolato, III= Prolato esferoidal.; IAP= Índice de Área Polar; GE= Grosor de la Exina; OE= Ornamentación de la exina, I= Microreticulado, II= Reticulado; LC= Longitud del colpo; AC= Ancho del colpo; NA= Número de apertura; CA= Característica de la Apertura, I= Sin vestíbulo, II= Con vestíbulo; LE= Longitud de la Endoapertura; AE= Ancho de Endoapertura; TE= Tipo de Endoapertura I= Lalongadas, II = Circulares; Todas las mediciones son en micrómetros =µm
| Caracteres/ Especies | EP | EE | R P/E | Forma | IAP | GE | OE | LC | AC | NA | CA | LE | AE | TE |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N. emarginata | 12.97±0.85 | 9.76±1.49 | 1.36 | II | 0.24 | 0.90±0.17 | I | 4.30±0.80 | 1.27±0.34 | 3 | I | 4.85±0.98 | 1.32±0.43 | I |
| N. hondurensis | 15.34±1.45 | 11.76±1.64 | 1.32 | I | 0.19 | 0.90±0.15 | I | 5.62±1.92 | 1.20±0.27 | 3 | II | 7.60±2.47 | 1.59±0.34 | I |
| N. paniculata | 9.81±2.87 | 7.41±2.25 | 1.34 | II | 0.21 | 0.84±0.20 | I | 3.45±0.66 | 1.12±0.21 | 3 | I | 5.26±1.57 | 1.25±0.24 | I |
| P. cordifolius | 12.34±1.11 | 11.15±1.16 | 1.11 | III | 0.22 | 1.19±0.22 | II | 4.79±0.92 | 1.52±0.44 | 3 | I | 6.61±0.93 | 1.18±0.43 | I |
| P. guatemalensis | 15.97±1.29 | 12.30±0.58 | 1.3 | I | 0.24 | 1.08±0.01 | I | 6.12±1.43 | 2.37±0.51 | 3 | II | 7.54±1.83 | 1.69±0.19 | I |
| P. mexicanaus | 17.13±1.23 | 13.82±1.62 | 1.25 | I | 0.27 | 1.12±0.17 | I | 5.55±0.64 | 2.49±0.94 | 3-4 | I | 5.61±0.81 | 5.61±0.81 | II |
Discusión
En este trabajo se exploran los patrones generales de la morfología polínica de Neomillspaughia y Podopterus así como también se registra su variación interespecífica con el propósito de reconocer su utilidad taxonómica. Como resultado de las observaciones los caracteres tales como la forma, número de aberturas, ornamentación y tipo de endoapertura de los granos de polen muestran variación a diferentes niveles taxonómicos. Las formas del grano de polen registradas en este estudio son de tres tipos: subprolato, prolato y prolato esferoidal. El primer tipo se presenta en Neomillspaughia hondurensis,
Podopterus guatemalensis y P. mexicanus, mientras que el segundo tipo lo exhibe N. emarginata y N. paniculata y en tercer tipo P. cordifolius. Las discrepancias en el tamaño y forma del grano de polen pueden reflejar variación intraespecífica (Oswald et al., 2011), diferentes estados de madurez del polen (García et al., 2015), deberse a diferencias entre material vivo y de herbario (Oswald et al., 2011), la elección en los métodos de extracción e incrustación (Ayodele, 2005) o al grado de hidratación de los granos de polen antes del proceso de fijación (Russell, 2003). De esta manera, tal vez se explique el por qué P. mexicanus en nuestro estudio, aparece con granos subprolatos y discrepa de Alfaro-Bates et al. (2010), que los describen como oblatos esferoidales. Las mediciones promedio de los ejes polar y ecuatorial de ambos autores es de P= 37.4 y E= 39.1 Las mediciones promedio obtenidas en nuestro estudio fueron de P=17.13 y E=13.82.
Especies estrechamente relacionadas a menudo producen granos de polen similares (Wang y Dobritsa, 2018). El polen para las tres especies que conforman el género Neomillspaughia se describe como prolato a subprolato con índice de área polar pequeña y ornamentación microrreticulada y es concordante con las observaciones generales realizadas por Nowicke y Skvrla (1977), Sánchez-Dzib et al. (2009) y Alfaro-Bates et al. (2010) en Neomillspaughia emarginata.
En cuanto al número de aperturas, el grano de polen en esta tribu es tricolporado (rara vez 4-colporado) y coincide con el tipo básico de polen en la familia Polygonaceae (Nowicke y Skvrla, 1977; Wodehouse, 1931). El número de aperturas también es importante en la separación de P. mexicanus de las otras especies de la tribu ya que esta especie puede presentar granos tri y tetracolporados en proporciones variables. Alfaro-Bates et al. (2010) indican que la proporción de granos tricolporados observados en P. mexicanus fue la misma para granos tetracolporados. En este estudio, la proporción de granos con cuatro aperturas fue menor. De acuerdo con García et al. (2015) esta tendencia que exhiben algunas plantas a incrementar el número de aperturas o a presentar granos con diferente número de aperturas en un mismo individuo, se debe a menudo a procesos de hibridación. También, se ha documentado que la variación en el número de aperturas está ligado al nivel de ploidía en los individuos (Albert et al., 2018; Reeder et al., 2016; Yurtseva et al., 2014). Por ejemplo, en Rumex acetosella se observó que la tasa de granos tricolporados es de 100% para individuos diploides y que esta disminuye al aumentar el nivel de ploidía tetraploide y hexaploide (Den Nijis et al., 1980). Asimismo, polen con diferente número de aperturas ha sido observado en algunas especies del género Polygonum y se ha asociado al nivel ploidal (Yurtseva et al., 2014). Podopterus mexicanus es también la única especie que presenta endoaperturas de tipo lolongado dentro de la tribu. En relación con la presencia de vestíbulo en las aperturas solo Podopterus guatemalensis y Neomillspaughia hondurensis lo poseen sin haberse observado en las otras cuatro especies.
La ornamentación es un carácter de suma importancia para reconocer a los géneros e incluso algunas especies (Hong et al., 2005; Wang y Dobritsa, 2018). En Neomillspaughia y Podopterus, la ornamentación más común es de tipo microrreticulada, únicamente en P. cordifolius es reticulada. Las ornamentaciones observadas en este estudio para los tres géneros son consistentes con lo observado por Wodehouse (1931), Nowicke y Skvrla (1977) y Brandbyge (1990a).
Los granos de polen en Podopterus cordifolius son los únicos en exhibir una forma prolato esferoidal y una exina reticulada con lúmenes profundos y bien definidos observables en microscopía óptica y de barrido. Esta especie presenta unas pocas poblaciones documentadas en Oaxaca y Colima y además difiere con las otras especies de Podopterus por poseer una ocrea larga y hialina, como las encontradas en los integrantes de la subfamilia Polygonoideae, presentar un hábito de crecimiento de tipo liana y flores de color lila, un carácter único en Polygonaceae (Burke et al., 2010). Si bien los estudios filogenéticos moleculares de la subfamilia Eriogonoideae (Burke et al., 2010; Koenemann y Burke, 2020) en los que se incluye a P. mexicanus y P. cordifolius considera a este género monofilético resultan contrastantes con la información palinólógica aquí presentada. En este contexto valdría la pena considerar mayor número de muestras en ambos estudios e incluir P. guatemalensis para corroborar la monofilia del género y para explicar las relaciones infragenéricas.
Conclusiones
La caracterización palinológica de las especies de Neomillspaughia y Podopterus ha permitido explorar la utilidad taxonómica en el contexto de la subfamilia Eriogonoideae corroborando que el número de aperturas y los patrones de ornamentación de la exina son similares a los encontrados en otros géneros. La variación en la forma, número y tipo de aperturas, así como también en la ornamentación de la exina de los granos de polen en las seis especies estudiadas son críticas para su identificación por lo que pueden ser útiles en estudios de otra índole como los melisopalinológicos. Neomillspaughia emarginata y Podopterus mexicanus especies de importancia de apícola y marcadores geográficos que contribuyen de manera importante en la producción de miel en la península de Yucatán (México).

