











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
El tomate (Solanum lycopersicum L.) es una de las hortalizas que más se consumen en el mundo (FAO, 2020). La popularidad de su consumo se debe a que se emplea en diferentes guisos y platillos, ya sea en estado fresco o industrializado. Con el incremento poblacional en las siguientes décadas habrá mayor demanda de productos alimenticios frescos o procesados (Melotto et al., 2020), lo que generará mayor economía en la producción y comercialización de productos primarios de origen vegetal, y se vería reflejado en aumento en las fuentes de trabajo.
En México, en los diferentes sistemas de producción con mayor superficie cultivada de tomate se emplean variedades mejoradas importadas, por lo que existe un alto grado de dependencia del germoplasma foráneo, lo que propicia fuga de divisas (Salgado-Meraz et al., 2018).
En varias regiones de México aún persisten y se manejan poblaciones nativas de especies cultivadas en sistemas tradicionales, el tomate es uno de los más notorios (Chávez et al., 2011). Se trata de un cultivo que se ha adaptado a diversas condiciones agroclimáticas y sistemas de producción, lo cual se ha demostrado en varios estudios; asimismo, se ha reportado la existencia de especies silvestres (Álvarez-Hernández et al., 2009; Chávez et al., 2011) y poblaciones nativas (Bonilla-Barrientos et al., 2014) en la República Mexicana, que tienen potencial como fuente de variabilidad genética.
A pesar de la disponibilidad de germoplasma, el mejoramiento genético de tomate en México no ha sido atendido de manera sistemática, ni por instituciones del gobierno ni por centros públicos de investigación (Salgado-Meraz et al., 2018), siendo así que la generación tanto de híbridos como de variedades mejoradas de tomate en el país es nula (SNICS, 2020).
El mejoramiento genético de cualquier especie vegetal debe partir de una amplia base genética; ésta se encuentra en diferentes fuentes y una de las más importantes son las poblaciones nativas y los parientes silvestres (Marín-Montes et al., 2020). Diversos autores han señalado que el tomate presenta limitantes en su proceso de mejora debido a la reducida variación genética que posee (Marín-Montes et al., 2019; Salgado-Meraz et al., 2018); ésto representa un riesgo mayor porque la ganancia genética en los ciclos de mejoramiento subsecuentes sería muy reducida. Para ampliar la base genética existen diversas estrategias, entre las que destacan la recolecta de germoplasma nativo y silvestre (Marín-Montes et al., 2019), e incluso el uso de mutágenos.
El manejo de los materiales nativos de tomate por agricultores ha conducido a la acumulación de genes que controlan diversos caracteres de interés antropocéntrico (Carrillo-Rodríguez et al., 2019); en cambio, durante el proceso de generación de variedades comerciales se fueron eliminando genes, de tal forma que su variabilidad genética se ha reducido, lo que ha impactado negativamente en la calidad funcional y nutraceútica del tomate y otras especies (Ebert, 2020). La actual preferencia de los consumidores por alimentos orgánicos, nutraceúticos, de mayor calidad y libres de residuos químicos es una condición que debe aplicarse en la mejora genética del tomate. En este sentido, en tomate se debe iniciar el proceso de mejora genética con la recolección de germoplasma, ya sea silvestre o nativo. El valor de la colección radica en el conocimiento específico de sus características fenotípicas y genéticas, mismas que requieren conocerse de manera detallada mediante evaluaciones de tipo agronómico, morfológico, molecular y genómico.
A México no se le considera centro de origen del tomate, pero si como el espacio geográfico de domesticación y diversificación (Peralta y Spooner, 2007), por lo que se infiere que posee amplia variabilidad genética (Blanca et al., 2012), razón por la cual se llevó a cabo este estudio con el objetivo de evaluar la diversidad morfológica y agronómica de 20 accesiones de tomate nativo del sur de México e identificar las promisorias para usarlas en mejoramiento genético y promover su conservación.
MATERIALES Y MÉTODOS
Sitio experimental
La investigación se llevó a cabo en el Campo Experimental Zacatepec del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), ubicado en Zacatepec de Hidalgo, Morelos, México, en las coordenadas 18° 39’ 16’’ N y 99° 11’ 54’’ O, a una altitud de 910 msnm, con clima cálido subhúmedo, lluvias en verano, precipitación anual promedio de 800 mm y temperatura anual promedio de 24 °C (García, 1981).
Material genético
Se evaluaron 20 poblaciones de tomate provenientes de los estados de Campeche (3), Morelos (1), Oaxaca (3), Puebla (9), Tabasco (1) y Veracruz (3), México (Cuadro 1). Las semillas de las poblaciones fueron recolectadas en diferentes años y conservadas a temperatura de 5 ºC.
Cuadro 1 Origen de las poblaciones de tomate nativo evaluadas en el ciclo otoño-invierno 2019. Zacatepec, Morelos, México.
| Clave | Forma de fruto | Municipio | Estado |
| JCM01 | Riñón | Dzitbalché | Campeche |
| JCM02 | Riñón | Huauchinango | Puebla |
| JCM03 | Riñón | Tlacolula | Oaxaca |
| JCM04 | Riñón | Poza Riza | Veracruz |
| JCM05 | Cherry grande | Xoxocotla | Morelos |
| JCM06 | Cherry | Tlacolula | Oaxaca |
| JCM07 | Riñón | Teapa | Tabasco |
| JCM08 | Medio riñón | Dzitbalché | Campeche |
| JCM09 | Medio Saladete | Tlacolula | Oaxaca |
| JCM10 | Riñón | Huauchinango | Puebla |
| JCM11 | Riñón | Xitlala | Puebla |
| JCM12 | Riñón | Zozocolco de Hidalgo | Veracruz |
| JCM13 | Cherry | Zozocolco de Hidalgo | Veracruz |
| JCM14 | Chino criollo | Altepexi | Puebla |
| JCM15 | Chino criollo | Altepexi | Puebla |
| JCM16 | Chino criollo | San Sebastián Zinacatepec | Puebla |
| JCM17 | Chino criollo | San José Miahuatlán | Puebla |
| JCM18 | Chino criollo | San Sebastián, Tehuacán | Puebla |
| JCM19 | Chino criollo | Tlacolitros, Zinacatepec | Puebla |
| JCM20 | Riñón | Dzitbalché | Campeche |
Manejo experimental de las poblaciones
Las 20 poblaciones de tomate, que fueron consideradas como los tratamientos, se establecieron en un invernadero modificado con techo plástico y paredes laterales cubiertas por malla, con temperatura promedio de 20 °C y humedad relativa de 79.1 %, durante el ciclo otoño-invierno (septiembre a diciembre) de 2019, bajo un diseño experimental completamente al azar con tres repeticiones, cada unidad experimental constó de ocho plantas establecidas en macetas de plástico de 30.48 cm de diámetro y 24 cm de altura.
La siembra se realizó el 3 de septiembre de 2019 en charolas de poliestireno de 50 cavidades con sustrato comercial Sunshine Mix® No. 3. En cada cavidad se depositaron dos semillas y se colocaron bajo condiciones ambientales de 26 °C y 70 % de humedad relativa durante cuatro días. Cuando la planta presentó la cuarta hoja verdadera, 28 días después de la siembra, se trasplantó en macetas con sustrato a base de ocochal (hojarasca de ocote descompuesta), atocle (suelo de vega de río) y polvillo de coco en block – Pelemix® en proporción 60:20:20 v/v/v. La planta se formó y tutoreó a un solo tallo con rafia para soporte en la etapa de crecimiento vegetativo y fructificación. El control de mosca blanca se realizó con la aplicación de 1.5 g L-1 de Confidor® (Imidacloprid) y de 4 g L-1 de sulfato de cobre contra tizón, ambos en forma preventiva. El riego de las plantas fue con un sistema por goteo y se aplicó solución nutritiva a base de nitrato de potasio (0.22 g L-1), fosfato monopotásico (0.21 g L-1), Kelatex® (0.032 g L-1) y ácido nítrico (0.25 g L-1).
Caracterización morfológica
En cinco plantas con competencia completa en cada unidad experimental se midieron la longitud del primer entrenudo (cm), la distancia de la base de la planta al primer racimo (cm), la distancia del primero al segundo racimo (cm), la distancia del segundo al tercer racimo (cm) y el diámetro de tallo (mm). Se contaron el número de nudos de la base de la planta al primer racimo, número de inflorescencias, número de flores por inflorescencia y número de frutos por planta. En flores del segundo racimo se midió la longitud de estambre (mm), de pétalo (mm), sépalo (mm) y pedicelo (mm). En todos los frutos de los tres primeros racimos se midió el ancho (cm), la longitud (cm) y se registró el peso (g) en una balanza digital (Ohaus®, Parsippany, New Jersey, EUA), con este valor de peso se estimó el rendimiento de fruto por planta. Para determinar la precocidad de las accesiones se contabilizaron los días transcurridos desde el trasplante hasta que se alcanzó 50 % de la floración del primer racimo.
Análisis estadístico
La información se analizó mediante análisis de varianza. Para la comparación de medias, para separar a las accesiones, se aplicó la prueba de Tukey (P ≤ 0.05). Con los promedios estandarizados de las variables evaluadas se realizó análisis de componentes principales y de conglomerados usando el método de Ward. El número de grupos formados en el dendrograma se determinó con la pseudoestadística t2 de Hotelling (Johnson, 2000) y se confirmó la diferencia estadística con el análisis de varianza. Todos los análisis se llevaron a cabo con el paquete estadístico SAS Versión 8 (SAS Institute, 2000).
RESULTADOS Y DISCUSIÓN
Análisis de varianza
Con el análisis de varianza se detectaron diferencias estadísticas significativas entre las poblaciones en todas las variables registradas, excepto en el diámetro de tallo. El coeficiente de variación fue de 1.43 a 33 %; los valores más altos se presentaron en el número de flores por inflorescencia, rendimiento de frutos por planta y longitud del primer entrenudo (Cuadro 2); ésto indicó la variación fenotípica presente en las poblaciones de tomate evaluadas. Valores altos de coeficientes de variación en número de flores por racimo, peso medio de frutos, frutos por racimo y total de frutos al quinto racimo fueron reportados por Carrillo y Chávez (2010) en accesiones del estado de Oaxaca. En otro estudio con 40 colectas de tomate nativo de los estados de Oaxaca y Puebla se reportó el mayor coeficiente de variación de 39.7 % en número de lóculos (Bonilla-Barrientos et al., 2014). En estas investigaciones los autores señalaron que la variación genética representa la materia prima para la mejora genética de la especie.
Cuadro 2 Cuadrados medios, promedios y coeficiente de variación de variables evaluadas en 20 poblaciones de tomate nativo en el ciclo otoño-invierno de 2019. Campo Experimental Zacatepec, Morelos, México.
| Variable | CM | Promedio | CV (%) |
| Longitud primer entrenudo (cm) | 3.35* | 5.68 | 23.28 |
| Altura de la base al primer racimo (cm) | 210.84** | 55.72 | 6.32 |
| Distancia del 1er racimo al 2do racimo (cm) | 37.52** | 24.52 | 10.98 |
| Distancia del 2do racimo al 3er racimo (cm) | 54.20** | 24.58 | 13.77 |
| Diámetro de tallo (mm) | 1.30NS | 10.93 | 8.14 |
| Nudos de la base al 1er racimo | 2.18** | 10.36 | 9.04 |
| Días a 50 % de floración del primer racimo | 5.45** | 53.90 | 1.43 |
| Número de inflorescencias | 4.51** | 6.64 | 13.35 |
| Número de flores por inflorescencia | 59.55** | 12.35 | 33.09 |
| Longitud del estambre (mm) | 1.82** | 8.23 | 5.74 |
| Longitud del pétalo (mm) | 12.24** | 16.75 | 10.82 |
| Longitud del sépalo (mm) | 47.46** | 12.51 | 11.43 |
| Longitud de pedicelo (mm) | 8.21** | 6.65 | 10.14 |
| Peso promedio del fruto (g) | 5227** | 65.46 | 17.31 |
| Longitud promedio del fruto (cm) | 782** | 3.97 | 6.24 |
| Ancho promedio del fruto (cm) | 495** | 4.40 | 6.34 |
| Número de frutos por planta | 58.06** | 15.95 | 18.81 |
| Rendimiento de fruto por planta (g) | 792665** | 922.9 | 24.65 |
CM: cuadrados medios, CV: coeficiente de variación, *: significativo con P ≤ 0.05., **: significativo con P ≤ 0.01, NS: no signifativo.
La mayor cantidad de inflorescencias la presentó la población JCM06, estadísticamente similar a otras tres poblaciones. En el número de flores por inflorescencia, el mayor promedio se obtuvo en la población JCM01, estadísticamente igual que otras 11 poblaciones (Cuadro 3).
Cuadro 3 Comparación de medias de siete caracteres de inflorescencia y fruto en poblaciones de tomate nativo en el ciclo otoño-invierno de 2019. Campo Experimental Zacatepec, Morelos, México.
| Población | NI | NFI | PF (g) | LF (cm) | AF (cm) | NFP | RTO (g) |
| JCM01 | 7.0 bcd | 22.2 a | 57.1 c | 3.0 def | 4.7 bcd | 16.8 a-d | 1017 b-g |
| JCM02 | 6.5 bcd | 18.4 ab | 44.7 cd | 3.3 b-f | 4.1 de | 21.0 ab | 934 c-h |
| JCM03 | 6.1 bcd | 14.5 ab | 37.8 cde | 3.0 c-f | 3.9 de | 21.8 ab | 738 e-i |
| JCM04 | 6.0 bcd | 18.2 ab | 42.0 cd | 3.3 b-f | 4.0 de | 21.0 ab | 900 d-h |
| JCM05 | 6.2 bcd | 14.8 ab | 62.7 bc | 3.5 bcd | 4.6 cd | 7.5 e | 436 g-j |
| JCM06 | 10.0 a | 9.5 b | 2.0 f | 1.4 g | 1.4 g | 16.3 a-e | 33 j |
| JCM07 | 6.4 bcd | 16.2 ab | 42.4 cd | 3.2 b-f | 4.1 de | 14.1 a-e | 597 f-j |
| JCM08 | 6.2 bcd | 7.1 b | 95.3 ab | 3.9 b | 5.8 a | 13.2 b-e | 1282 a-f |
| JCM09 | 7.1 bcd | 9.0 b | 17.6 def | 2.7 ef | 2.9 f | 15.8 a-e | 294 hij |
| JCM10 | 6.5 bcd | 6.4 b | 102.9 a | 3.8 bc | 5.8 a | 8.4 de | 896 d-h |
| JCM11 | 8.3 abc | 11.4 ab | 21.41 def | 2.7 ef | 3.2 ef | 18.4 abc | 392 g-j |
| JCM12 | 7.7 abcd | 18.0 ab | 38.3 cde | 2.6 f | 4.1 d | 21.0 ab | 824 d-h |
| JCM13 | 8.6 ab | 8.2 b | 3.7 ef | 1.7 g | 1.7 g | 23.3 a | 87 ij |
| JCM14 | 5.6 cd | 9.4 b | 111.8 a | 6.0 a | 5.3 abc | 12.5 b-e | 1418 a-e |
| JCM15 | 5.5 d | 8.9 b | 107.2 a | 5.9 a | 5.2 abc | 11.5 cde | 1279 a-f |
| JCM16 | 5.4 d | 9.3 b | 120.3 a | 6.2 a | 5.5 ab | 13.2 b-e | 1614 abc |
| JCM17 | 5.2 d | 9.8 ab | 112.2 a | 6.2 a | 5.5 ab | 12.8 b-e | 1450 a-d |
| JCM18 | 5.7 cd | 10.8 ab | 121.0 a | 6.3 a | 5.6 a | 16.3 a-e | 1664 ab |
| JCM19 | 5.4 d | 9.9 ab | 121.4 a | 6.2 a | 5.5 ab | 14.5 a-e | 1726 a |
| JCM20 | 6.8 bcd | 14.4 ab | 46.47 cd | 3.4 bcde | 4.2 d | 18.3 abc | 868 d-h |
NI: número de inflorescencias, NFI: número de flores por inflorescencia, PF: peso de fruto, LF: longitud de fruto, AF: ancho de fruto, NFP: número de frutos por planta, RTO: rendimiento de fruto por planta. Medias con la misma letra en las columnas son estadísticamente iguales (Tukey, P ≤ 0.05).
En peso de fruto, siete poblaciones mostraron superioridad (Cuadro 3). En comparación con lo reportado por Carrillo-Rodríguez et al. (2019), los promedios obtenidos en el presente estudio fueron inferiores, estos autores reportaron peso de fruto de 192 g. El mayor tamaño de fruto en términos de longitud fue obtenido en seis poblaciones (Cuadro 3). Estos mismos autores reportaron la mayor longitud de fruto en poblaciones nativas de Oaxaca (5.47 cm), que es inferior a lo obtenido en el presente estudio, donde el mayor promedio fue de 6.3 cm.
Las poblaciones JCM08, JCM10 y JCM18 presentaron el mayor ancho de fruto, pero fueron estadísticamente similares a otras cinco poblaciones (Cuadro 3). Estos resultados son inferiores a los reportados por Carrillo-Rodríguez et al. (2019) en poblaciones nativas de Oaxaca, donde el mayor ancho registrado fue de 9.7 cm. La mayor cantidad de frutos por planta se obtuvo en la población JCM13, que es un material silvestre del estado de Veracruz (Cuadro 3); este resultado coincide con lo obtenido por Salgado-Meraz et al. (2018), quienes mencionaron que esta característica favorable es distintiva de las poblaciones silvestres.
El mayor rendimiento de fruto por planta se alcanzó en la población JCM19, que fue estadísticamente igual a otras seis poblaciones (Cuadro 3). Salgado-Meraz et al. (2018) reportaron mayor rendimiento de fruto, 2453 g por planta, muy superior a lo obtenido en el presente estudio; asimismo, Carrillo-Rodríguez et al. (2019) observaron un rendimiento de fruto por planta de 2059 g. En otros estudios con poblaciones nativas de tomate, Marín-Montes et al. (2019) reportaron los mayores promedios con valores de 6.39 cm de longitud de fruto, 5.74 cm en ancho de fruto, peso promedio de fruto de 83.14 g y rendimiento de fruto por planta de 257 g. Por su parte, las poblaciones JCM06 y JCM13 mostraron los menores promedios en número de flores por inflorescencia, peso, tamaño y rendimiento de fruto por planta (Cuadro 3).
Análisis de componentes principales
En el análisis multivariado de componentes principales (CP) se encontró que el 82.35 % de la variación morfo-agronómica total se explicó con los tres primeros componentes. Las variables con mayor valor descriptivo de la variabilidad en el CP1 fueron características relacionadas con estructuras florales, tamaño y peso de fruto, así como el rendimiento por planta; en el CP2 influyeron principalmente longitud de nudos y altura de la planta hasta el primer racimo y en el CP3 la precocidad de los materiales (Cuadro 4). La variación explicada por los tres primeros componentes en este estudio fue similar a lo reportado por Carrillo y Chávez (2010) en materiales de Oaxaca con 86.25 %. Marín-Montes et al. (2019) en un estudio con 26 accesiones de tomate provenientes de los estados de Puebla, Oaxaca y Veracruz indicaron que la variación fenotípica explicada con los tres primeros componentes fue de 81.5 %. En otro estudio con 40 accesiones de tomates nativos de los estados de Oaxaca y Puebla, México se encontró que los dos primeros componentes principales explicaron 77.03 % de la variación total (Bonilla-Barrientos et al., 2014). En todos estos estudios los caracteres con mayor valor descriptivo fueron similares y se refieren a tamaño de planta, floración, tamaño de fruto y rendimiento de fruto por planta.
Cuadro 4 Variables de mayor valor descriptivo, varianza acumulada, vectores y valores propios en el análisis de componentes principales (CP) de tomate nativo en el ciclo otoño-invierno de 2019. Campo Experimental Zacatepec, Morelos, México.
| Variables | CP1 | CP2 | CP3 |
| Longitud primer entrenudo (cm) | -0.0008 | 0.4258 | 0.0615 |
| Altura de la base al primer racimo (cm) | 0.1366 | 0.4055 | -0.0234 |
| Nudos de la base al 1er racimo | 0.0973 | 0.4308 | -0.2394 |
| Días a 50 % de floración del primer racimo | 0.1974 | 0.0035 | -0.5972 |
| Número de inflorescencias | -0.2929 | 0.0140 | 0.4119 |
| Longitud del estambre (mm) | 0.3032 | -0.0208 | 0.1500 |
| Longitud del pétalo (mm) | 0.3066 | -0.1013 | 0.1306 |
| Longitud del sépalo (mm) | 0.3065 | -0.1594 | 0.0278 |
| Peso promedio del fruto (g) | 0.3141 | -0.1292 | 0.0204 |
| Longitud promedio del fruto (cm) | 0.3015 | -0.0297 | -0.1351 |
| Ancho promedio del fruto (cm) | 0.3117 | -0.0266 | 0.0588 |
| Rendimiento de fruto por planta (g) | 0.3117 | -0.0266 | 0.0588 |
| Valores propios | 8.92 | 3.97 | 1.09 |
| Varianza explicada (%) | 52.50 | 23.38 | 6.46 |
| Varianza acumulada (%) | 52.50 | 75.88 | 82.35 |
La representación de la diversidad se muestra en la Figura 1, empleando los dos primeros componentes principales. En el cuadrante IV se agruparon siete poblaciones que se caracterizaron por mayor peso, tamaño y rendimiento de fruto por planta, así como mayor tamaño de estructuras florales. En el cuadrante III se ubicaron dos grupos, uno formado por JCM06 y JCM13, los cuales se caracterizaron por menor tamaño, peso y rendimiento de fruto, mientras que el otro fue conformado por JCM10 con mayor peso y ancho de fruto. En el centro de los cuadrantes se concentraron cinco poblaciones con promedios intermedios en tamaño y rendimiento de fruto por planta. Por último, en la parte superior del cuadrante II se localizaron cinco poblaciones con la mayor altura de la base de la planta al primer racimo, mayor longitud y cantidad de nudos al primer racimo.

Figura 1 Dispersión de 20 poblaciones nativas de tomate sobre el plano determinado por los primeros dos componentes principales. Ciclo otoño-invierno de 2019, Campo Experimental Zacatepec, Morelos, México.
Con base en análisis de componentes principales, Bonilla-Barrientos et al. (2014) identificaron cuatro grupos de diversidad; en cambio, Sanjuan-Lara et al. (2014) reportaron seis grupos en 120 familias derivadas de una población nativa. Las diferencias con este trabajo se explican por los patrones propios de variación genética del germoplasma evaluado.
Análisis de conglomerados
En este análisis se encontraron dos grandes grupos, uno conformado por tres subgrupos y el otro por dos. El análisis de varianza respectivo determinó que los cinco grupos del conglomerado (Figura 2) son estadísticamente diferentes (P ≤ 0,01).
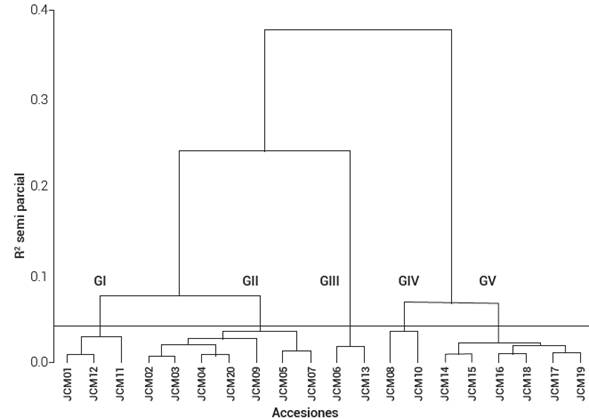
Figura 2 Dendrograma de 20 poblaciones de tomate nativo evaluadas en el ciclo otoño-invierno de 2019. Campo Experimental Zacatepec, Morelos, México.
El Grupo 1, integrado por tres accesiones, se caracterizó por tener el mayor valor en los caracteres de altura de la base al primer racimo, distancia entre los racimos 2 y 3, distancia entre los nudos, número de inflorescencias y flores por inflorescencia. El Grupo 2, conformado por siete accesiones, presentó la segunda mayor distancia de la base de la planta al primer racimo, en la cantidad de nudos, número de flores por inflorescencia, y el segundo más tardío en días a floración. El Grupo 3 aglomeró a dos accesiones que sobresalieron en ancho de fruto, pero con el menor número de flores por inflorescencia; el segundo con la mayor longitud de estambre, longitud de pétalo, longitud de sépalo y longitud de pedicelo; también el segundo en peso, largo de fruto y rendimiento de fruto por planta. Al Grupo 4 lo integraron dos accesiones tipo cereza, que se caracterizaron por presentar los promedios más bajos en caracteres de fruto, estructura de la flor, rendimiento, y fueron precoces; sin embargo, produjeron gran cantidad de frutos. El Grupo 5 integró a seis accesiones, presentó los mayores promedios en estructura de la flor, largo de fruto, peso de fruto, rendimiento de fruto por planta, fueron tardíos y tuvieron el menor número de inflorescencias (Cuadro 5).
Cuadro 5 Valores promedio de variables en cinco grupos formados con el análisis de conglomerados de poblaciones de tomate nativo en el ciclo otoño-invierno de 2019. Campo Experimental Zacatepec, Morelos, México.
| Variables | GI | GII | GIII | GIV | GV |
| Longitud primer entrenudo (cm) | 6.98 | 5.85 | 5.65 | 4.65 | 5.20 |
| Altura de la base al primer racimo (cm) | 65.52 | 57.69 | 50.95 | 38.57 | 55.82 |
| Distancia del 1er al 2do racimo (cm) | 28.87 | 22.70 | 23.89 | 18.53 | 26.69 |
| Distancia del 2do al 3er racimo (cm) | 30.61 | 24.11 | 21.80 | 17.27 | 25.50 |
| Nudos de la base al 1er racimo | 11.1 | 10.8 | 9.8 | 8.6 | 10.1 |
| Días a 50 % de floración del primer racimo | 52 | 54 | 53 | 51 | 54 |
| Número de inflorescencias | 7.7 | 6.4 | 6.4 | 9.3 | 5.5 |
| Número de flores por inflorescencia | 17.2 | 15.1 | 6.7 | 8.9 | 9.7 |
| Longitud del estambre (mm) | 8.13 | 8.01 | 8.60 | 6.54 | 8.98 |
| Longitud del pétalo (mm) | 16.12 | 16.05 | 16.64 | 13.42 | 19.06 |
| Longitud del sépalo (mm) | 10.68 | 10.75 | 11.03 | 7.05 | 17.82 |
| Longitud de pedicelo (mm) | 6.13 | 5.80 | 6.13 | 4.68 | 8.76 |
| Peso promedio del fruto (g) | 38.98 | 42.00 | 99.13 | 2.93 | 115.70 |
| Longitud promedio del fruto (cm) | 2.80 | 3.27 | 3.88 | 1.57 | 6.20 |
| Ancho promedio del fruto (cm) | 4.06 | 4.00 | 5.84 | 1.60 | 5.49 |
| Número de frutos por planta | 18.7 | 17.1 | 10.8 | 19.8 | 13.6 |
| Rendimiento de fruto por planta (g) | 744.73 | 681.34 | 1089.48 | 60.57 | 1525.78 |
Los análisis estadísticos univariado y multivariado confirmaron la existencia de una amplia variación morfológica en las poblaciones de tomate evaluados en este estudio. Esta variación coincide con lo obtenido en 49 muestras poblacionales de tomates silvestres y semidomesticadas procedentes del estado de Oaxaca (Carrillo y Chávez, 2010). También, son similares a los resultados obtenidos en la evaluación morfológica y agronómica de muestras procedentes del Centro y Sureste de México, donde se identificaron materiales locales con alta variabilidad fenotípica (Vásquez-Ortiz et al., 2010); asimismo, permitió la identificación de poblaciones con características sobresalientes en tamaño, peso y rendimiento de fruto por planta, como lo señalan Bonilla-Barrientos et al. (2014), los cuales podrían utilizarse directamente como variedades para los mercados locales y regionales, o como fuente de genes para el mejoramiento genético, que aún no se aborda de manera sistematizada en México.
CONCLUSIONES
En la muestra de poblaciones nativas de tomate estudiadas la variación existente se basa principalmente en caracteres relacionados con la planta, precocidad, tamaño de fruto y rendimiento. Las variables con mayor valor descriptivo de la variabilidad fueron estructuras florales, tamaño y peso de fruto, rendimiento por planta, longitud de nudos y altura de la planta hasta el primer racimo y precocidad. Se determinó la estructura de agrupamiento de las accesiones estudiadas en dos grandes grupos, uno conformado por tres subgrupos y el otro por dos. Se identificaron seis poblaciones con los mayores promedios en estructura de la flor, largo de fruto, peso de fruto y rendimiento de fruto por planta.

