










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkActa zoológica mexicana
versión On-line ISSN 2448-8445versión impresa ISSN 0065-1737
Acta Zool. Mex vol.20 no.1 Xalapa abr. 2004
Detección de poblaciones de Rhagoletis pomonella (Diptera: Tephritidae) durante la fructificación de Crataegus mexicana (Rosaceae) en Puebla, México
Vicente Hernández-Ortiz1, Irma Morales2, y Carlos Vergara3
1 Instituto de Ecología A.C., Departamento de Entomología, Ap. Postal 63. Km 2.5 antigua carretera a Coatepec No. 351, Congr. El Haya, CP 91070, Xalapa, Veracruz, MÉXICO. hernanvi@ecologia.edu.mx.
2 Benemérita Universidad Autónoma de Puebla, Escuela de Biología. Cd. Universitaria, CP 72570, Puebla, Puebla, MÉXICO.
3 Universidad de las Américas, Puebla, Departamento de Química y Biología. Santa Catarina Mártir, CP 72820, Cholula, Puebla, MÉXICO.
Recibido: 29 de abril 2003
Aceptado: 3 de octubre 2003
RESUMEN
Durante el período de fructificación-maduración del tejocote (Crataegus mexicana), se realizó una evaluación de las poblaciones de Rhagoletis pomonella (Walsh), con el propósito de detectar las etapas críticas de la infestación. El estudio se llevó a cabo en el Municipio de Huejotzingo, Puebla, para lo cual se seleccionaron cuatro localidades de muestreo. La detección de adultos se efectuó empleando 64 trampas tipo McPhail durante 15 semanas (entre septiembre y diciembre 1997), distribuidas entre las cuatro localidades, y para estimar los niveles poblacionales de infestación, fueron muestreados 1680 frutos maduros durante todo el estudio. Los resultados mostraron que la mayor captura de adultos se presentó desde los inicios de septiembre hasta principios de octubre (durante las primeras cinco semanas del estudio), cuando la mayoría de los frutos habían alcanzado su desarrollo e iniciaban su maduración. Las pruebas estadísticas aplicadas no mostraron diferencias significativas de captura entre los cuatro sitios de muestreo (F(2,42)= 1.58, P= 0.21), pero en cuanto a la distribución temporal (semanas), el efecto si fue significativo (F(14,42)= 5.53, P< 0.001). La captura total entre las trampas mostró que las poblaciones de adultos tienen una distribución agregada, debido a que solo algunas trampas en cada localidad obtuvieron los mayores porcentajes de captura. Con base en los resultados obtenidos, se analizan y discuten las etapas de mayor susceptibilidad del tejocote a la infestación de frutos por R. pomonella en la región.
Palabras clave: Rhagoletis pomonella, poblaciones, infestación, Crataegus mexicana.
ABSTRACT
An assessment of the Rhagoletis pomonella populations was made to detect the critical period of infestation of the "tejocote" (Crataegus mexicana) during its fruiting and maturation. The study was conducted in four localities in Huejotzingo Municipality, Puebla. Adult populations were monitored by use of 64 McPhail traps distributed among the four localities during 15 weeks (from September to December 1997), while estimation of infestation levels in fruits were estimated by sampling 1,680 mature fruits throughout the study. Results showed that most captures of adults were from the early part of September to the beginning of October (during the first five weeks of the study), when most fruits had attained their full development and ripeness was initiated. Statistical results showed no significant differences of captures among sampled sites (F(2,42)= 1.58, P= 0.21), unless respect to temporal distribution (weeks), difference of significance was observed (F(14,42) = 5.53, P< 0.001). The total of captures among traps showed that adult populations had an aggregate distribution, because only some traps at each locality accounted for most of the captures. On the basis of our results, we discuss the time periods of higher susceptibility of the tejocote to infestation by R. pomonella in this region.
Key words: Rhagoletis pomonella, populations, infestation, Crataegus mexicana.
INTRODUCCIÓN
El grupo de especies pomonella comprende cuatro especies descritas: R. pomonella (Walsh), R. mendax Curran, R. zephyria Snow, y R. cornivora Bush, las cuales se distribuyen particularmente en Norteamérica, no obstante, al menos la primera mantiene poblaciones alopátricas en México (Bush 1966). Varios estudios han señalado que diversas poblaciones de R. pomonella en Norteamérica, que se alimentan de hospederos distintos presentan diferencias genéticas, por lo que ésta especie se reconoce en la actualidad como un complejo de especies crípticas (McPheron et al. 1988, Berlocher et al. 1993, Berlocher 2000). En este sentido, el estado taxonómico de algunas poblaciones mexicanas que infestan especies de Crataegus es incierto, puesto que difieren de las demás especies del grupo pomonella en el locus mitocondrial COII, lo cual sugiere que estas poblaciones también podrían constituir una especie distinta (Smith & Bush 1997).
Debido a la utilización de diferentes plantas hospederas por especies estrechamente relacionadas del grupo pomonella que habitan en una misma área, han sido objeto de estudio para proponer la formación de razas hospederas mediante procesos de especiación simpátrica, cuyos principales argumentos estarían dados por cambios en el uso de hospederos relacionados y diferencias en sus periodos de fructificación (Bush 1969, Bush 1975). Rhagoletis pomonella se encuentra asociada principalmente con algunos grupos de la familia Rosaceae, y en Norteamérica se reconocen alrededor de 15 especies de hospederos de Crataegus, en tanto que para México apenas se conocen C. mexicana Moc. et Sessé y Malus pumila Miller (Bush 1966, Smith & Bush 2000).
En México, el género Crataegus L. se distribuye principalmente entre los 1000 y 2800 m de altitud, en regiones donde los niveles de precipitación pluvial se estiman entre los 500 y 1500 mm anuales ocupadas por bosques de Pino-Encino (Rzedowski 1988). De acuerdo con la revisión taxonómica de Crataegus (Phipps 1997), en nuestro país han sido descritas 13 especies de tejocotes, de las cuales solo C. mexicana se encuentra en el estado de Puebla, conocida también con otros nombres como C. pubescens (H.B.K.) Steud., C. subserrata Benth. y C. hypolasia K. Koch, que en la actualidad se consideran como sinonimias.
Para la detección de las poblaciones de Rhagoletis han sido utilizadas diversos tipos de trampas a través de estímulos visuales como formas y colores, olores sintéticos de frutos, y atrayentes alimenticios (Duan & Prokopy 1992, Frías et al. 1993, Reynolds et al. 1996, Rull & Prokopy 2000). En Chile, las poblaciones de R. tomatis Foote asociadas con Lycopersicon esculentum Miller, fueron evaluadas mediante trampas rectangulares y esféricas, además de estímulos visuales (color rojo y amarillo) y alimenticios (proteína hidrolizada), cuyos resultados mostraron que las esferas amarillas obtuvieron la mayor eficiencia de captura (Frías et al. 1993). La evaluación de poblaciones de R. mendax Curran, que infestan especies de Vaccinium en Norteamérica, demostró que el uso de trampas esféricas rojas y verdes resultaron más eficientes en la captura de moscas sexualmente inmaduras, comparadas con la trampa Pherocon AM, por lo cual proponen la utilización de las trampas esféricas de color, en etapas tempranas de la fructificación cuando aun no existen individuos sexualmente maduros (Teixeira & Polavarapu 2001). En tanto que los resultados de un estudio enfocado a evaluar la eficiencia de atracción en diferentes estados fisiológicos de R. pomonella en huertos de manzana, basado en la comparación de trampas cebadas con olores atrayentes basados en frutos y la presencia-ausencia de alimento, sugieren que las estrategias de trampeo deben enfocarse al uso de atrayentes basados en olores de frutos, en lugar de atrayentes alimenticios (Rull & Prokopy 2000).
En México, el tejocote tiene una producción nacional estimada en 28,000 toneladas anuales, cultivado principalmente en los estados de México, Michoacán y Puebla que aportan el 60% de la producción (INEGI 1990). No obstante, pese a las pérdidas económicas atribuidas a la R. pomonella, no existe un protocolo regular para evaluar sus poblaciones, o en su caso, se utilizan los mismos protocolos (tanto biológicos como de detección) establecidos para otras moscas de la fruta (i.e., Anastrepha spp.), por lo cual no existen estudios documentados de su biología y relaciones fenológicas con sus plantas hospederas. En virtud de los daños ocasionados por esta especie en huertos de la región occidental de Puebla, y debido a la carencia de recursos económicos de los productores para implementar medidas distintas a las tradicionalmente empleadas, se determinó realizar un estudio de detección poblacional de R. pomonella basado en el uso de trampas tipo McPhail, usando un atrayente alimenticio y agregando un estimulo visual, que nos permitiera conocer las etapas críticas de la infestación de C. mexicana en la región.
MATERIALES Y MÉTODOS
Región de estudio. El estudio se llevó a cabo en el municipio de Huejotzingo, en el Oeste de Puebla, Mexico, localizado en las cercanías del Volcán Popocatépetl (19°14' 19°16'N; 98°20' - 98°38' W) a una elevación de 2,260 m sobre el nivel del mar (INEGI 1993). Las comunidades de vegetación nativa de la región han sido reemplazadas en su mayoría por cultivos en los valles, pero en las zonas montañosas, se presentan bosques de Pino-Encino asociados con vegetación arbustiva, entre las especies dominantes se encuentran Pinus montezumae, P. teocote y P. oocarpa, además de algunas especies de encinos como Quercus laurina y Q. affinis (Soto & Aguiar 1969). El clima de la zona es templado subhúmedo con estación lluviosa en el verano entre abril y octubre (García 1981), con una temperatura anual promedio de 15.7°C, y una precipitación pluvial anual de 868.2 mm; la temperatura varía de 12.4°C en enero hasta 18.2°C en mayo, en tanto que la precipitación mínima es de 4.9 mm en marzo, y la máxima de 195.6 mm durante julio (Fig. 1). La localidad de estudio incluye huertos mixtos compuestos principalmente de tejocote (Crataegus mexicana), nogal (Juglans mollis) y capulín (Prunus serotina), además de algunos árboles de manzana (Malus domestica).
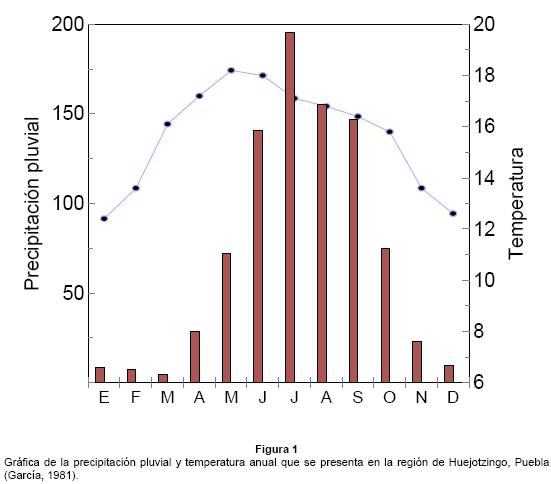
La fenología de Crataegus mexicana en esta región inicia con la floración entre febrero y marzo, los primeros brotes aparecen entre marzo y abril, y el desarrollo de los frutos se extiende hacia finales de septiembre; la maduración ocurre entre octubre y noviembre, y el periodo de cosecha se inicia alrededor de mediados de noviembre hasta diciembre (Deolarte, com. personal).
Para la evaluación de las poblaciones se establecieron cuatro sitios de muestreo en las siguientes localidades: Sitio A) San Bartolo; Sitio B) Mextla; Sitio C) Coyuco; y Sitio D) Ciprés. En estas localidades, los productores realizaron aplicaciones de insecticida para reducir el riesgo de infestación mediante aplicaciones mensuales irregulares desde inicios de la floración. En San Bartolo se realizaron cinco aplicaciones (marzo, mayo, junio, julio y agosto); en Mextla, se efectuaron solo dos aplicaciones (marzo y julio); en Coyuco cuatro (marzo, junio, julio, y primera semana de septiembre) y en Ciprés también se realizaron cuatro aplicaciones (abril, mayo, julio y tercera semana de septiembre).
Detección de adultos. En cada localidad se instalaron 16 trampas tipo McPhail, por lo que en total se emplearon 64 trampas en toda la región en una proporción de aproximadamente dos trampas por hectárea. Las trampas se pintaron de color amarillo intenso en su mitad inferior, y se utilizó como atrayente una mezcla de 235 ml de agua, 13 ml de proteína hidrolizada (Nulure, de Bayer) y 5 gr de borato de sodio (bórax) como conservador. Estas se colocaron aproximadamente a 2/3 de la altura del árbol, y la distancia entre ellas fue de 40-50 m dependiendo de la distancia entre los árboles del huerto. El atrayente fue reemplazado cada siete días durante las 15 semanas del estudio entre septiembre y diciembre de 1997. Los adultos capturados se lavaron y preservaron en alcohol 70%.
Revisión de frutos. Para evaluar los niveles de infestación natural en cada sitio, adicionalmente se recolectaron muestras semanales de frutos maduros tanto del árbol como del suelo dependiendo de su disponibilidad. Durante las primeras seis semanas del estudio, la disponibilidad de frutos maduros fue prácticamente nula; a partir del 3 de noviembre (semana N9) alrededor del 50% de los frutos presentaron una coloración amarilla y se consideró como su estado de madurez. Desde esa fecha hasta el final del estudio se recolectaron semanalmente 60 frutos maduros, seleccionados principalmente de cuatro árboles en donde se colocaron trampas en cada sitio, haciendo un total de 420 frutos por sitio, y obteniendo un total de 1,680 frutos para los cuatro sitios durante el estudio. Los frutos fueron colocados en recipientes de plástico con arena como sustrato de pupación y se mantuvieron en condiciones medioambientales de alrededor de 20-24° C. El número de pupas recuperadas se contabilizó semanalmente para cada uno de los sitios.
Análisis de datos. Para determinar los niveles de la variación poblacional de cada semana, se calculó el índice MTD en cada una de las localidades usando la siguiente fórmula: MTD= M/(TxD), en donde: M= número de moscas capturadas; T= número de trampas utilizadas; D= número de días de exposición de la trampa.
Para determinar las posibles diferencias de captura entre los sitios de colecta y entre las semanas, se aplicó un ANOVA paramétrico de dos vías, en donde el índice MTD fue transformado a arco seno para cumplir con los supuestos de normalidad y homocedasticidad (Zar 1984). Todos los análisis estadísticos se ejecutaron mediante el programa Statistica (1999). Para estimar la distribución espacial de los adultos se calculó el índice de dispersión obteniendo la varianza/ media muestral ID= S2/·· (Southwood 1978).
RESULTADOS
La comparación del total de moscas capturadas entre los cuatro sitios indicó que la mayor incidencia de adultos se registró en el Ciprés (36.6%), seguido de San Bartolo (33.4%), mientras que en Mextla y Coyuco, las proporciones fueron menores con el 20.1% y 12.9% respectivamente. En este sentido, los análisis estadísticos no mostraron diferencias significativas de captura entre los cuatro sitios de colecta (F(2,42)= 1.58, P= 0.21).
Respecto a la distribución temporal (semanas), el efecto de captura si fue significativo (F(14,42)= 5.53, P< 0.001), reflejado en que las poblaciones de adultos se detectaron principalmente durante las cinco primeras semanas del estudio (desde principios de septiembre al 6 de octubre), en este periodo se capturó el 90.5% de las moscas en los cuatro sitios, en tanto que entre las semanas O6 - N10 (7 de octubre al 10 de noviembre) se capturó el 8.7%, mientras que durante las últimas cinco semanas (N11-D15), solo se registró el 0.8% de las moscas.
En la localidad de San Bartolo, la mayor incidencia de captura se observó en las semanas S2 (MTD= 0.40) y S4 (MTD= 0.29); en Mextla ocurrió en las semanas S2 (MTD= 0.15) y O5 (MTD= 0.29); en Coyuco solo se presentó un valor elevado en la semana S4 (MTD= 0.38); y en Ciprés se presentaron las mayores incidencias de captura en las semanas S2 (MTD= 0.45) y S4 (MTD= 0.26) (Fig. 2).
En términos generales, para la región de Huejotzingo se observaron dos fuertes incrementos de la abundancia de adultos durante las semanas S2 y S4, y posteriormente a esas fechas, las poblaciones decrecen notablemente hasta desaparecer después de la primera semana de diciembre (Fig. 3).
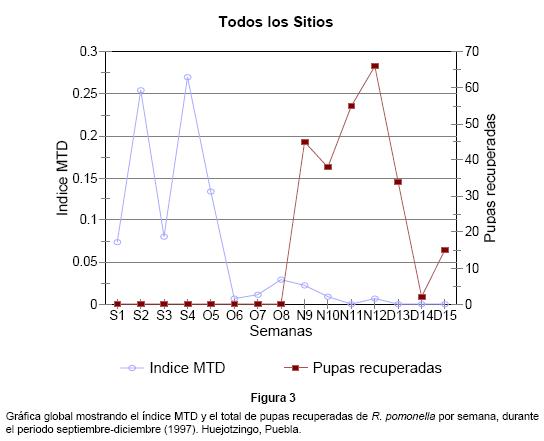
En relación con la infestación de frutos evaluada a través de las pupas recuperadas, los resultados generales mostraron que las infestaciones más elevadas en todas las localidades se presentaron desde la primera semana de noviembre (N9) hasta la primera semana de diciembre (D13), pero en toda la región es evidente que los mayores porcentajes de infestación se presentaron en las semanas N9 y N12. En este sentido, la localidad de Coyuco, registró un solo incremento durante la semana N12, lo cual coincide con un solo incremento de la población de adultos. De acuerdo con estos resultados, desde el inicio de la revisión de frutos maduros a partir de la semana N9 ya se encontraban larvas en cantidades importantes, por lo que es probable que desde principios de octubre ya se encuentren frutos infestados en bajas proporciones.
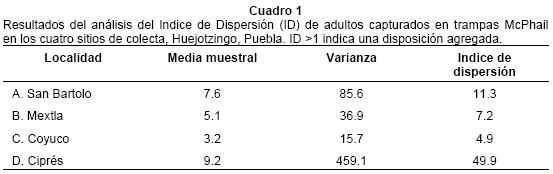
El porcentaje de capturas registradas en cada trampa fue muy variable, puesto que en una misma localidad se presentaron trampas que no capturaron ningún individuo, en tanto que otras capturaron las mayores proporciones. En San Bartolo la trampa A9 registró el 25% de capturas; en Mextla, la trampa B24 registró el 30%; en Coyuco la trampa C44 poco más del 30%; y en Ciprés la trampa D64 registró casi el 60% de las capturas (Fig. 4). Con base en la captura de las trampas, se analizó la distribución espacial de los individuos en cada localidad, calculando el índice de dispersión. Estos resultados indicaron que existe una fuerte agregación en todos los sitios estudiados (Cuadro 1).
DISCUSIÓN
En la localidad Coyuco (sitio C) se obtuvo la captura más baja, en tanto que en el Ciprés (sitio D) fue la más alta, esta diferencia podría deberse al efecto producido por los tratamientos químicos realizados por los productores, en particular la última aplicación realizada en Coyuco (durante la primera semana de septiembre), la cual coincidió con la aparición de poblaciones de adultos en toda la región, por lo que en ésta localidad se detectó un solo pico poblacional durante la última semana de septiembre (S4).
De acuerdo con la mayor incidencia de captura de adultos, concentrada principalmente entre las semanas S2 y O5, sugiere que existe una relación directa con las épocas del desarrollo de frutos, debido a que las observaciones de campo señalaron que a principios de septiembre se alcanza el máximo de frutos desarrollados en las cuatro localidades, mientras que la maduración (considerada como la presencia de frutos amarillos) tuvo sus inicios alrededor de la quinta semana (O5).
La presencia de dos picos poblacionales tanto de adultos como de larvas, sugiere una gran sincronización de la emergencia de adultos en relación con el proceso de desarrollo y maduración de frutos. De acuerdo con Webster y Stoffolano (1970), la distribución y abundancia de sus sustratos de alimentación constituyen un papel fundamental en la determinación de sus características de desarrollo, longevidad y fecundidad. Por ello, es probable que el periodo observado entre los dos mayores picos poblacionales de adultos (S2 y S4) e infestaciones registradas (N9 y N12), corresponden a una generación que transcurre en ocho semanas, por lo cual el ciclo de vida desde la oviposición (determinada mediante la presencia de adultos) hasta el desarrollo de larvas maduras (determinada por la presencia de pupas recuperadas) transcurre en aproximadamente 56 días. Estos datos concuerdan con la información previamente registrada para esta especie, la cual se desarrolla entre 37 y 59 días desde su periodo de pre-oviposición hasta la formación de la pupa (Christenson & Foote 1960).
La alta disponibilidad de recursos predice una amplia distribución para optimizar el uso de los mismos, o bien que la presencia de enemigos naturales inducen ciertos patrones de distribución agregada (Williams et al., 2001). En este sentido, los estudios realizados en huertos de manzana en los EUA, se observó que en las poblaciones de R. pomonella, tanto larvas como pupas tienden a concentrarse en áreas donde las manzanas se encuentran agregadas, lo cual les confiere una mayor tasa de sobrevivencia a factores como la desecación y depredación (Cameron & Morrison 1974). Sin embargo, el fenómeno de agregación resultado de las capturas en trampas, podría tener sustento en la dependencia de un conjunto de factores ecológicos y fisiológicos relacionados con estímulos visuales y olfatorios para localizar sitios de alimentación y oviposición (Aluja & Prokopy 1993).
En términos de las medidas que deben implementarse para el control de la R. pomonella en la región de Huejotzingo, es necesario considerar que los adultos aparecen poco tiempo después de que se alcanzó la máxima precipitación pluvial en la región (julio), mientras que el decremento poblacional se acentúa cuando la temperatura media mensual y la precipitación pluvial inician su descenso (octubre). Basados en estos argumentos, y en el hecho de que las poblaciones de R. pomonella tienen una diapausa en el estado pupal, resulta innecesaria la aplicación de tratamientos desde la floración enfocados a disminuir el daño ocasionado por esta especie. Por ello se recomienda que la detección de adultos se inicie desde mediados del mes de agosto, cuando probablemente emergen los primeros individuos de la generación. Los resultados obtenidos sobre el incremento de la población de adultos, sugieren que la etapa más susceptible de infestación del tejocote en la región, ocurre en un periodo de aproximadamente seis semanas, la cual tiene sus inicios alrededor de la tercera o cuarta semana de agosto, y se extiende hasta finales de septiembre o primera semana de octubre.
AGRADECIMIENTOS
Nuestro agradecimiento a la "Unión de Fruticultores Huexotl" de la región de Huejotzingo, en especial a los señores: Celerino Deolarte, Raúl Morales, Manuel Maretano y Rodolfo Morales, por todas las facilidades y apoyo que nos brindaron durante el desarrollo del trabajo de campo. De igual forma, agradecemos a José A. Gómez Anaya (Departamento de Entomología, INECOL) por su ayuda durante la revisión de datos y análisis estadísticos de los resultados obtenidos. Esta investigación constituye una contribución al proyecto "Insectos fitófagos y saprófagos", que se desarrolla en el Departamento de Entomología, del Instituto de Ecología A.C.
LITERATURA CITADA
Aluja, M. & R. J. Prokopy. 1993. Host odor and visual stimulus interaction during intratree host finding behavior of Rhagoletis pomonella flies. J. Chem. Ecol. 19: 2671-2696. [ Links ]
Berlocher, S. H. 2000. Radiation and divergence in the Rhagoletis pomonella species group: inferences from allozymes. Evolution 54: 543-557. [ Links ]
Berlocher, S. H., B. A. McPheron, J. L. Feder & G. L. Bush. 1993. Genetic differentiation at allozyme loci in the Rhagoletis pomonella (Diptera: Tephritidae) species complex. Ann. Entomol. Soc. Am. 86: 716-727. [ Links ]
Bush, G. L. 1966. The taxonomy, cytology and evolution of the genus Rhagoletis in North America (Diptera: Tephritidae). Bull. Mus. Comp. Zool. 134: 438-443. [ Links ]
----------. 1969. Sympatric host race formation and speciation in frugivorous flies of the genus Rhagoletis (Diptera: Tephritidae). Evolution 23: 237-251. [ Links ]
----------. 1975. Sympatric speciation in phytophagous parasitic insects, pp: 187-206. In: P.W. Pierce, Ed. Evolutionary strategies of parasitic insects and mites,. Plenum, New York, USA. [ Links ]
Cameron, P. J. & F. O. Morrison. 1974. Sampling methods for estimating the abundance and distribution of all life stages of the apple maggot, Rhagoletis pomonella (Diptera: Tephritidae). Can. Entomol. 106: 1025-1034. [ Links ]
Christenson, L. D. & R. H. Foote. 1960. Biology of fruit flies. Ann. Rev. Entomol. 5: 171-192. [ Links ]
Duan, J. J. & R. J. Prokopy. 1992. Visual and odor stimuli influencing effectiveness of sticky spheres for trapping apple maggot flies Rhagoletis pomonella (Walsh) (Dipt., Tephritidae). J. Appl. Entomol. 113: 271-279. [ Links ]
Frías, D. L., C. González, A. Henry & A. Alviña. 1993. Distribución geográfica y respuesta visual de Rhagoletis tomatis Foote (Diptera: Tephritidae) a trampas esféricas y rectángulos de diferentes colores. Acta Ent. Chilena 18: 185-194. [ Links ]
García, E. 1981. Modificaciones al sistema de clasificación climática de Köppen. Instituto de Geografía, Univ. Nac. Aut. Mex., México, 164 pp. [ Links ]
INEGI. 1990. Anuario estadístico de los Estados Unidos Mexicanos. Instituto Nacional de Estadística Geografía e Informática, México. [ Links ]
----------. 1993. Huejotzingo, Estado de Puebla. Cuaderno Estadístico Municipal. Instituto Nacional de Estadística Geografía e Informática, México. [ Links ]
McPheron, B. A., D. C. Smith & S. H. Berlocher. 1988. Genetic differences between host races of Rhagoletis pomonella. Nature 336 (6194): 64-66. [ Links ]
Phipps, J. B. 1997. Monograph of Northern Mexican Crataegus (Rosaceae, Subfam. Maloideae). Sida, Botanical Miscellany 15: 1-94. [ Links ]
Reynolds, A. H., R. J. Prokopy, T. A. Green & S. E. Wright. 1996. Apple maggot fly (Diptera: Tephritidae) response to perforated red spheres. Florida Entomol. 79: 173-179. [ Links ]
Rull, J. & R. J. Prokopy. 2000. Attraction of apple maggot flies, Rhagoletis pomonella (Diptera: Tephritidae) of different physiological states to odour-baited traps in the presence and absence of food. Bull. Entomol. Res. 90: 77-88. [ Links ]
Rzedowski, J. 1988. Vegetación de México. Edit. Limusa, Cuarta reimpresión. México, 432 pp. [ Links ]
Smith, J. J. & G. L. Bush. 1997. Phylogeny of the genus Rhagoletis (Diptera: Tephritidae) inferred from DNA sequences of mitochondrial cytochrome oxidase II. Mol. Phylog. Evol. 7: 33-43. [ Links ]
----------. 2000. Phylogeny of the subtribe Carpomyina (Trypetinae), emphasizing relationships of the genus Rhagoletis, pp: 187-217. In: M. Aluja and A.L. Norrbom, Eds. Fruit flies (Tephritidae): phylogeny and evolution of behavior. CRC Press, Boca Raton, USA. [ Links ]
Soto, M. C. & L. F. Aguiar. 1969. El uso del suelo en la región de Huejotzingo y San Martín Texmelucan, Puebla, México. Univ. Nac. Aut. Mex., México, pp: 12-22. [ Links ]
Southwood, T. R. E. 1978. Ecological methods with particular reference to the study of insect populations. Chapman and Hall. London, England, pp: 27-44. [ Links ]
Statistica, 1999. Software for PC statistical analysis. Statsoft Inc., Tulsa, Oklahoma. [ Links ]
Teixeira, L. A. F. & S. Polavarapu. 2001. Effect of sex, reproductive maturity stage and trap placement, on attraction of the blueberry maggot fly (Diptera: Tephritidae) to sphere and Pherocon AM traps. Florida Entomol. 84: 363-369. [ Links ]
Webster, R. P. & J. G. Stoffolano. 1970. The influence of diet of the reproductive system of the apple maggot, Rhagoletis pomonella. Ann. Entomol. Soc. Am. 71: 844-849. [ Links ]
Williams, I. S., T. H. Jones & S. E. Hartley. 2001. The role of resources and natural enemies in determining the distribution of an insect herbivore population. Ecol. Entomol. 26: 204-211. [ Links ]
Zar, J. H. 1984. Biostatistical analysis. Second Edition. Prentice-Hall, Inc. New Jersey, USA, 718 pp. [ Links ]

