











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
The number of monarch butterflies (Danaus plexippus L.) has decreased due to three main factors: degradation in hibernation zones in Mexico; reduction in breeding areas in the United States due to the loss of cottonwood plants (Asclepias spp.; host plants), as well as land-use change; and changes in climatic conditions (Brower et al., 2012). Reforestation of the Monarch Butterfly Biosphere Reserve (MBBR), on the borders of the states of Michoacán and Mexico, is important to increase the number of trees where the butterfly overwinters.
Abies religiosa (Kunth) Schltdl. & Cham. is the dominant forest species at overwintering sites. Based on our direct experience, nursery plant production of this species entails greater difficulty than plant production of the genus Pinus, due to the following reasons: (a) the female cones of A. religiosa are produced almost exclusively at the upper end of the crown, which makes collection difficult and expensive (Arriola et al., 2015; Nieto de Pascual-Pola, Musálem, & Ortega-Alcalá, 2003), especially when they are large trees; (b) A. religiosa usually has a high percentage of empty seeds, a low germination percentage (Iglesias-Andreu et al., 2010; Nieto de Pascual-Pola et al., 2003) and a short life under storage conditions (Arriola et al., 2015; Carrillo, Patiño, & Talavera, 1980; Nieto de Pascual-Pola et al., 2003; Salazar & Soihet, 2001); and (c) obtaining permits for the collection of A. religiosa seed in the MBBR’s core area is complicated. This last difficulty occurs because rule 62 of the MBBR Management Program (Comisión Nacional de Áreas Naturales Protegidas [CONANP], 2001) stipulates that during the monarch butterfly hibernation season, a period that coincides with the ripening of cones in November and December, it is prohibited to engage in activities that put the butterfly at risk. Additionally, when the seed is collected by personnel external to the communities and ejidos of the MBBR, a permit is required from SEMARNAT (Secretaría del Medio Ambiente y Recursos Naturales, 2018), which, in order to grant it, requests that the applicant demonstrate the consent of the communities (SEMARNAT-02-004 procedure). The consent document is prepared for each of the communities and ejidos in which seed is collected. This process is complex, since it is difficult to obtain the minimum legal quorum of the ejidal and communal assemblies, when the motive is only to give permission.
This situation has led to the production of nursery plants using seed from outside the MBBR’s core area, which implies that the plant does not adapt due to a lack of coupling between the climate of the seed collection site and the planting site (Castellanos-Acuña et al., 2018; Castellanos-Acuña, Lindig-Cisneros, Silva-Farías, & Sáenz-Romero, 2014; Johnson, Sorensen, St. Clair, & Cronn, 2004). As an alternative, the ejidatarios (ejido members) and communeros (joint land holders) of the MBBR have raised the possibility of rescuing naturally regenerated plants that, based on their practical experience, would otherwise die in the dry season following their germination. A thick layer of moss (>10 cm) is common in the area, which acts as a mechanical barrier, preventing the seedlings that germinate and emerge from reaching the soil (Rodríguez-Laguna, Razo-Zárate, Fonseca-González, Capulín-Grande, & Goche-Telles, 2015). In many cases, the seedlings do not survive the dry season of the following year (Madrigal-Sánchez, 1967; Manzanilla, 1974; Marañón et al., 2004), because the moss dehydrates (Ángeles-Cervantes & López-Mata, 2009; Chen et al., 2015) or acts as a competitor for the seedling of this conifer (Stuiver, Wardle, Gundale, & Nilsson, 2014).
In a sampling of A. religiosa populations in an altitudinal transect (2 850 m to 3 350 m) on San Andrés hill, 50 km west of the MBBR, Castellanos-Acuña et al. (2014) observed an altitudinal pattern of morphological variation in needles and cones, where higher-altitude populations showed longer needles and shorter cones than low-altitude populations. Generally, an assay of A. religiosa provenances collected along an altitudinal gradient indicates an important genetic differentiation between populations, considering that such expression can be quantified whenever the plants grow in the same environment (Sáenz-Romero, Beaulieu, & Rehfeldt, 2011; Sáenz-Romero et al., 2012; Sáenz-Romero, Ruiz-Talonia, Beaulieu, Sánchez-Vargas, & Rehfeldt, 2011). Populations originating at lower altitudes produce plants with greater height, their dominant bud elongates more, they cease growth later in the year and they accumulate higher stem and foliage biomass, but are more susceptible to frost damage than plants originating at higher altitudes (Ortiz-Bibian et al., 2017); however, such a pattern has not been confirmed in provenance trials with seed collected directly from the MBBR.
Based on the above, the objective of the present study was to determine the viability of producing A. religiosa in a nursery from naturally regenerated seedlings, and the genetic variation of quantitative traits among populations, associated with an altitudinal gradient.
Materials and methods
The recently germinated natural regeneration seedling was collected in an altitudinal gradient (2 960 to 3 450 m) located within the MBBR in the Casablanca, Guadalupe Buenavista and La Mesa ejidos, municipality of San José del Rincón, State of Mexico (Table 1). The seedlings germinated in situ in late spring and at the beginning of the rainy season in 2015 and were collected between the last week of October and the first week of November of the same year. The selected seedlings correspond to sites where there is a thick layer of moss (> 5 cm, personal observation), which is produced by the large amount of moisture in these areas during the time when the natural regeneration of A. religiosa is established.
Table 1 Location of the collection sites (provenance) of Abies religiosa seedlings and of the nursery trial site in the municipality of San José del Rincón, State of Mexico.
| Provenance | Altitude (m) | N latitude | W longitude |
|---|---|---|---|
| Provenance 1 | 3 450 | 19° 34’ 05.4” | 100° 13’ 53.0” |
| Provenance 2 | 3 350 | 19° 34’ 28.0” | 100° 13’ 05.5” |
| Provenance 3 | 3 239 | 19° 34’ 44.2” | 100° 12’ 51.6” |
| Provenance 4 | 3 157 | 19° 35’ 24.0” | 100° 12’ 31.2” |
| Provenance 5 | 3 052 | 19° 35’ 44.5” | 100° 12’ 06.9” |
| Provenance 6 | 2 960 | 19° 36’ 56.0” | 100° 11’ 13.3” |
| Nursery trial site | |||
| Poblado La Mesa | 3 000 | 19° 35’ 18.2” | 100° 10’ 46.1” |
In each altitudinal site, the collection was done through a selection of seedlings still with a testa, cotyledonary leaves (without primary leaves), and a thin, pink and vigorous stem with a height less than 5 cm. The seedlings were moved from the field to the nursery in containers, where the roots with naturally attached soil were covered with an additional layer of local soil (Figure 1).

Figure 1 Transplanting of recent Abies religiosa seedlings that emerged naturally in the field into containers in a community forest nursery in the La Mesa ejido, San José del Rincón, State of Mexico. (a) Seedling naturally emerged at sites with a thick layer of diverse moss and mulch. (b) Extraction of natural regeneration plant. (c) Transport of seedling from the collection site to the nursery; the roots were protected by the use of moist local soil, in addition to the naturally adhered soil. (d) Transplant into nursery bag with local soil. (e) Provenance test; each nursery bed (three) contains two blocks (six in total).
At the test site, weeds were removed and the nursery was covered with a 50 % mesh shadehouse to prevent predation by animals, dehydration of plants by direct exposure to the sun, and damage by strong winds or hail. Immediately after their collection, the seedlings were transplanted into polyethylene bags (8 x 7 x 17 cm) for a nursery that contained local soil (previously sifted) as substrate from sites dominated by A. religiosa.
The experimental design was six completely randomized blocks. In total, there were 200 seedlings from each of the six provenances, randomly distributed within the blocks in continuous plots of 36 seedlings, arranged in three rows of 12 individuals, except block number six, in which there were only 20 seedlings per provenance. Seedlings were placed at the ends of the nursery beds to avoid the edge effect. The six blocks were placed in three beds (two blocks per bed) to facilitate the measurement work with accessible corridors.
The height from the seedling was assessed from the base to the tip of the main bud (mm) and the basal diameter (0.1 mm) of the stem were evaluated bimonthly and semiannually, respectively, for 18 months (December 2015 to May 2017), as well as the survival of each plant. The initial height or basal diameter was used as a covariate to rule out the effect of the initial size of the plant at the time of transplant to the nursery bag, because the exact germination date was not known.
An analysis of variance was performed using the SAS statistical package version 9.1 (SAS Institute, Inc. 2004) with the PROC GLM procedure, using the following statistical model:
where,
Y ijk |
observation of the k th individual in the j th provenance of the i-th block |
µ |
overall mean |
B i |
effect of the i th block |
P j |
effect of the j th provenance |
B i *P j |
interaction between block and provenance (source of variation used as the error term to determine the significance of block and provenance) |
S ijk |
effect of covariate due to initial plant size |
E ijk |
error |
Provenance and block were considered random effects; the same model was used for survival, but without a covariate. Additionally, a regression of means of the variables evaluated by provenance was made against the altitude of provenance, in order to determine the association between the growth potential and the altitudinal gradient.
Results
The average survival of A. religiosa seedlings was 75 % (Figure 2) 18 months after establishment in the nursery and approximately two years after the possible date of germination in the field. Table 2 indicates that the provenances caused significant differences in the survival (P < 0.0001), height (P = 0.0430) and basal diameter (P < 0.0001) of A. religiosa.
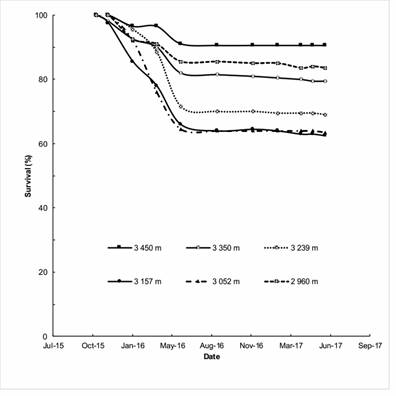
Figure 2 Survival curves of six provenances of naturally regenerated Abies religiosa plants in the Monarch Butterfly Biosphere Reserve.
Table 2 Analysis of variance of the growth variables of six provenances of Abies religiosa from the Monarch Butterfly Biosphere Reserve.
| Source of variation | df | Height | Basal diameter | Survival | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| MS | P | MS | P | MS | P | |||||
| Block | 5 | 4 469 | 0.3959 | ns | 80.4 | 0.5346 | ns | 0.53 | 0.1133 | ns |
| Provenance | 5 | 10 715 | 0.0430 | * | 732.6 | <0.0001 | *** | 2.26 | <0.0001 | *** |
| Block*Provenance | 25 | 4 227 | 0.0002 | *** | 97.5 | 0.0011 | ** | 0.26 | 0.2272 | ns |
| Covariate | 1 | 28 827 | <0.0001 | *** | 438 | 0.0021 | ** | --- | --- | |
| Error | 718 | 1 774 | 46 | 0.22 | ||||||
df = degrees of freedom (height and basal diameter = 718; survival = 1 164); MS = mean square. *P ≤ 0.05; **P ≤ 0.01; *** P ≤ 0.001; ns = not significant.
According to Figure 3 and Table 3, the provenance with the most live plants was the one with the highest altitude (3 450 m), being statistically similar to the lowest-altitude provenance (2 960 m), while the lowest survival was an intermediate-altitude provenance (3 157 m); regarding plant height, the 3 350 m provenance had the highest value and was statistically similar to the provenances of the altitudinal extremes.
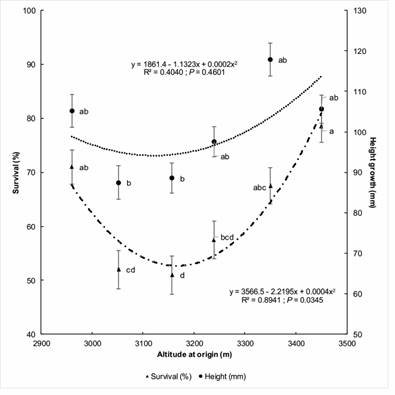
Figure 3 Quadratic regression of survival and final height of six Abies religiosa provenances against altitude of origin. The same letters indicate non-significant difference between provenances, according to Tukey's test (P ≤ 0.05). Vertical bars indicate the standard error of the mean.
Table 3 Comparison of means of growth variables of six provenances of Abies religiosa at 18 months of establishment in a nursery.
| Provenance | Altitude (m) | Height (mm) | Basal diameter (mm) | Survival (%) |
|---|---|---|---|---|
| 1 | 3 450 | 105.6 ab | 21.2 a | 78.5 a |
| 2 | 3 350 | 117.8 a | 20.1 ab | 67.5 abc |
| 3 | 3 239 | 97.6 ab | 16.7 bc | 57.5 bcd |
| 4 | 3 157 | 88.5 b | 15.6 c | 51 d |
| 5 | 3 052 | 87.5 b | 15.0 c | 52 cd |
| 6 | 2 960 | 105.1 ab | 18.3 abc | 71 ab |
| LSD | 25.6 | 3.9 | 16 |
Mean values with the same letters between provenances indicate non-significant difference (Tukey, P ≤ 0.05). LSD = least significant difference.
The altitudinal clinal pattern shown in Figure 4 indicates that the highest-altitude provenance had the greatest basal diameter and its value was statistically similar to that recorded in the 2 960 m (lowest altitude) and 3 350 m plants.

Discussion
The altitudinal clinal pattern of survival, height and basal diameter, in terms of the good performance of the higher-altitude provenances, is different from that reported for other conifers in Mexico, such as Pinus pseudostrobus Lindl. (Viveros-Viveros, Sáenz-Romero, López-Upton, & Vargas-Hernández, 2005) or Pinus hartwegii Lindl. (Loya-Rebollar et al., 2013). In these species, lower-altitude populations had larger basal diameters than higher-altitude ones, growing under optimal or suboptimal conditions. On the other hand, the data generated in the present study coincide with the good performance of early age (13-month-old) P. hartwegii seedlings, originating from seed of the populations of the upper altitudinal extreme (4 050 m) of Cofre de Perote and grown in a shadehouse in Xalapa, Veracruz (Viveros-Viveros, Marín-Hernández, Aparicio-Rentería, & Sáenz-Romero, 2018).
It is important to emphasize that the results of Ortiz-Bibian et al. (2017), in which less growth is reported in the higher-altitude provenances, represent the growth potential (the genetically conditioned capacity to grow under optimal conditions), while the conditions of the present work were much more difficult, since the nursery was a very basic facility and located at 3 000 m. Therefore, it is possible that a high provenance*environment interaction induces a different growth pattern. Additionally, it is necessary to evaluate the fact that the altitudinal pattern is not very pronounced (the difference between provenances of the altitudinal gradient is relatively minor), and that the last evaluation was made in May (in July they were planted in a field trial). There is a possibility that the pattern would have been different if the last evaluation had been made in November (2017), when the apical buds cease to grow and are at rest. Finally, the age of the plants can be considered too young to show a definite trend of their growth capacity in a nursery at 3 000 m. It is therefore recommended that these results be treated with caution.
It is also recommended to conduct further research on the use of naturally regenerated plants for growth in a nursery, for reforestation purposes, in order to avoid costs in the collection of seed directly from the standing tree.
Conclusions
A. religiosa plants from the altitudinal extremes had the highest average survival, height, and basal diameter values. The average survival of 75 % indicates that transplanting naturally regenerated plants, for later growth in a nursery, is viable. Therefore, the reintroduction in the field of the rescued naturally regenerated seedlings can be a complementary alternative to the usual reforestation programs, in order to achieve the ecological restoration of the overwintering A. religiosa stands of the Monarch butterfly.

