











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
Nearly fifty-six percent of the global human population lives in medium-sized cities harboring between 100,000 to 500,000 inhabitants. It has been predicted that, by the year 2035, 62.5 % of the human population will live in a city (UN-HABITAT 2020). Such an increase in the world’s urban population implies: 1) The establishment of new cities or the growth of those already established to the detriment of natural landscapes; since, this entails the extirpation of native flora and fauna at rates even higher than those produced by agriculture or forestry (Marzluff and Ewing 2001; McKinney 2006). 2) The establishment and increase of non-native wildlife species adapted to urban conditions and, eventually, the homogenization of the landscape (McKinney 2006).
The expansion of the urban territory onto the natural landscape alters its structure in terms of composition (cover classes) and configuration (spatial arrangement of cover classes; Liu et al. 2016), reconfiguring the landscape. Additionally, fragmented and disorderly growth (coupled with the lack of associated regulations) favors urban configurations where limits (or peripheries) are unclear. Since poorly urbanized neighborhoods are constantly added to the peripheries, they represent points of growth blurring the limits of cities (Terradas et al. 2011). This gives rise to cities with a broad variety of landscapes, with a varying degree of disturbance, sometimes mixed with fragments of conserved vegetation (Doygun 2009).
The reconfiguration of urban space alters the communities of native flora and fauna, in terms of species composition, abundance, richness and equitability (Aronson et al. 2014; Lättman et al. 2014). Wildlife has been reported to present different types of response to urbanization, according to the taxonomic group in question (Magle et al. 2019); for example, large predatory mammals are poorly adapted to survive in large cities (Ordiz et al. 2013). In contrast, other medium and large mammals, such as raccoons (Procyon lotor) or capybaras (Hydrochoerys hydrochaeris), are well adapted to urban conditions in some regions of the continent (Dybas 2017; Serra-Medeiros et al. 2021).
In the case of small mammals such as rodents, there are species that respond positively to urbanization, increasing their abundances (Santini et al. 2019); the so-called “urban dwellers”, which live permanently in the urban environment. The abundance increase of these species represents serious economic and public health problems for humans, similar those generated from the non-native rodent species like Mus musculus and Rattus species in the World (Meerburg et al. 2009; McFarlane et al. 2012). Other species exist that make occasional visits: “urban visitors” (Baker et al. 2003). Finally, some species respond negatively, reducing their abundances (Lopucki and Kitowski 2017), and are known as “urban avoiders” (McKinney 2006).
Previous studies have shown that urbanization influences the diversity of native rodent species, regardless of city size, reducing native species richness while increasing richness of non-native species. At the same time, these studies recognize the importance of preserving fragments of the landscape with natural coverage, in order to maintain high diversity of native rodents in urban areas (Rizkalla and Swihart 2009; Fernández and Simonetti 2013; Johnson and Karels 2016; Lopucki and Kitowski 2017). It is also important to control stray dogs and cats, common in many cities throughout the continent, which can be predators of native species and endanger their survival (Coronel-Arellano et al. 2021; Mella-Méndez et al. 2022).
The main objective of this work was to evaluate the response of the rodent community to the urbanization process in San Cristóbal de Las Casas, Chiapas, México (SCLC). For this purpose: 1) The alpha and beta diversity of the rodent community were recorded and compared against urbanization gradients; 2) the relationships between landscape metrics (in terms of configuration and composition) with rodent species richness and abundance were evaluated; and 3) based on the landscape-site approach, the study area was categorized into different categories of landscapes. We explain the rodent species distribution and suggest areas for conservation based on landscape categories and rodent diversity.
Materials and Methods
Study area. The city of San Cristóbal de Las Casas, Chiapas (SCLC; 16° 44’ 12” N, -92° 38’ 18” W) is in the region of The Highlands of Chiapas. The main ecosystem types in the city are: mountain wetlands (Ramsar sites; The Ramsar Convention Secretariat 2013), coniferous forests (pine, oak and pine-oak), and cloud forests (Trucíos-Caciano et al. 2013). The original layout of SCLC was made up of 18 blocks and 12 streets (Aubry 2008; Appendix 1). The city continued to expand towards the mountains, and towards the south of the valley, where the wetlands are located (Cantú Luna and Fenner Sánchez 2020). Today, it is a medium-sized city of 202,459 inhabitants (INEGI 2021). The urban area has increased by 12.5 % between 2001 and 2018 (Camacho-Valdez et al. 2019). Natural areas embedded in the city have reduced their size, and the current fragments have been designated for conservation in private, federal, and municipal reserves: Huitepec Ecological Reserve, Moxviquil Ecological Reserve, El Encuentro Ecological Park, Montetik Natural Park, La Kisst Wetland, and María Eugenia Wetland (Supplementary material 1).
Delimiting and characterizing the study area. The study area was delimited by 6 km radius circumference (Figure 1a). The characterization was conducted through a supervised classification of a Sentinel 2 image, of 10 m resolution. The image, from 2020, was analyzed using QGIS 3.16 (QGIS Development Team 2021) and the SCP (Semiautomatic Classification Plugin) plug-in 6.4 (Congedo 2020). The classification was performed with the method Maximum Similarity Algorithm. The accuracy of the classification was established by the Kappa coefficient at 80 % confidence value per land cover class. The area, was classified into four land cover classes, served for the subsequent categorization of the sampling sites: urban (soil covered by concrete, devoid of vegetation, houses, roads), forests (altered coniferous forest, oak forest, pine-oak forest), agriculture (agricultural plots, sown agricultural, agricultural in preparation, pastures), and wetlands (grasslands, areas covered by water).
Rodent species sampling. Seven bimonthly rodent samplings were performed, three in 2019 (June, August and October), one in 2020 (January) and three in 2021 (March, May and July), along seven urbanization gradients, established through the study area in straight line, and directed to the cardinal points (see Figure 1a). Along each of the seven urbanization gradients, nine sampling sites were established for each one: at 0 (1), 1.5 (2), 3.5 (3) and 6 km (3; Figure 1a). The minimum distance between sampling sites was 500 m. At each sampling site, 20 Sherman traps were placed simultaneously, for a total of 180 traps per gradient, per sampling month. The sampling sites had the following characteristics: at 0 km, the urban sites included houses, parking lots, hotels or hostels. Then, at 1.5 km, two contrasting sites were chosen, forests and wetlands or agricultural plots or urban areas. At 3.5 km, three contrasting sites were chosen, forests and wetlands and agricultural plots or pastures or urban areas. Finally, at 6 km, the selection of three contrasting sites was repeated.
In each sampling site, the traps were placed in two lines of 10 traps each one, with a separation of 10 meters between each trap and 10 meters between lines. However, in the case of traps placed in houses, five traps were located per house (totaling four houses). Traps were placed inside rooms, kitchens, grain storage warehouses, corrals, or patios. All traps were baited with a mixture of oats, peanut butter, and sunflower seeds. The traps were checked every morning at 7:00 hrs. Each specimen sampled was identified to species level with specialized guides and taxonomic keys (Álvarez-Castañeda et al. 2017); sex, reproductive condition and weight were recorded, in addition to conventional somatic measurements.
Rodents were marked with ear notches (Sántiz-Vázquez 2018) and released in situ. Each ear sample was stored in 70 % alcohol, for future genetic analysis. Two specimens of each native species by location were collected and deposited in the Mammal Collection of El Colegio de la Frontera Sur (ECOSUR). All non-native rodents were sacrificed and conserved in 70 % alcohol. The scientific collection license FAUT-0143 provided to CL from SEMARNAT was used. Rodents were handled according to the guidelines of the American Society of Mammalogists (Sikes and The Animal Care and Use Committee 2016).
Categorizing the study area. To categorize the study area into different landscape categories, we used a site-landscape approach (Fahrig 2013). This approach consisted of establishing a buffer of 500 m ratio in each effective sampling site, considering landscape units (LU; Figure 1b). The buffer was set to the home range of the rodent species previously collected in the region (San-José et al. 2014; Rodríguez Macedo 2016; Arce-Peña et al. 2019).
Each LU was characterized in terms of its composition and configuration, using eight landscape metrics obtained with FRAGSTATS 4.2.1 software (McGarigal and Ene 2015). We chose two metrics for composition: PLAND (percentage of landscape / land cover class), and Shannon’s diversity index (SHDI). The land cover classes considered were: urban (URB), forest (FOR), agricultural (AGRO), and wetland coverage (WET). Other six metrics for configuration were, number of patches (NP), contagion index (CONTAG), percentage of like adjacencies index (PLADJ), division index (DIVISION), patch richness (PR), and aggregation index (AI). The definition of each landscape metric is in Supplementary material 2.
Through a Principal Component Analysis (PCA), the spatial relationship between the LUs, the landscape metrics, and the relative abundance were explored via correlations. With the resultant ordination, landscape categories were assigned according to the structural characteristics with the highest loadings. The threshold to recognize those components best explaining variation was the asymptote of the distribution of the variance. This analysis was performed using the FactoMineR package (Le et al. 2020) in the R 4.10 programming language (R Core Team 2018).
Data Analysis. Alpha diversity was calculated using Hill numbers 0D (species richness), 1D (exponential of the Shannon diversity index), and 2D (inverse Simpson index; Moreno et al. 2011). Diversity analyzes were performed using the Vegan 1.8 package (Oksanen et al. 2007). Range-abundance graphs were built to simultaneously visualize abundance and diversity in the categories of LU (Magurran 1988). Differences in abundance and richness were estimated with a Hutcheson t-test (Zar 2010) between landscape categories. Additionally, beta diversity (i. e., species turnover) was calculated using the multiplicative-partition-of-diversity method. Furthermore, the Sorensen dissimilarity index was used to determine the dissimilarity between the LUs categories, as well as the partition of beta diversity into its components βSOR = βSIM + βSNE; for which we used the betapart 1.5.4 package (Baselga and Orme 2012). Finally, the hypothesis of a relationship between rodent diversity and landscape metrics was tested, using a series of generalized linear models with the Glmulti 1.0.8 package (Calcagno and de Mazancourt 2010).
Results
Characterization of the study area. The study area covered 15,143 ha; forests of oak, pine, pine-oak or oak-pine forests, in some state of conservation covered the 66 % of the area, urban 14 %, agricultural crops and pastures 18 %, and conserved mountain wetlands less than 2 % (Figure 1b).
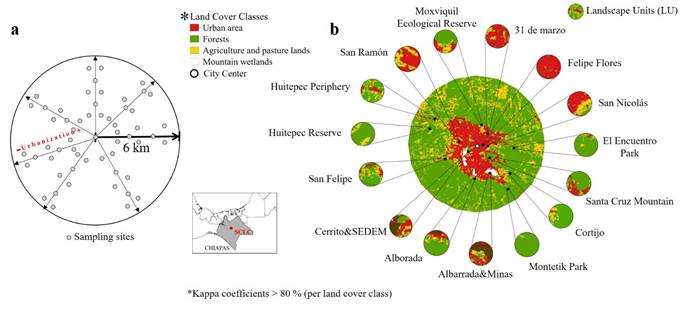
Figure 1 a) Sampling design for rodents in San Cristóbal de Las Casas, Chiapas, México. Arrows = representing urbanization gradients performed in this study; b) classified land use classes and landscape units (LU) in San Cristóbal de Las Casas, Chiapas, México.
Categorization of the study area. Rodents were captured only in 15 out of 63 sampling sites, such sites were established as landscape units (LUs; Figure 1b). Landscape metrics are presented in Table 1. The study area was categorized into four categories of landscape: urban, of transition, of mosaic and conserved. This categorization was conducted using principal component analysis (PCA). The first principal component (PC1) explained 53.1 % of the variation and was interpreted as a proxy for habitat heterogeneity (Figure 2), since it showed higher weights for the number of patches (NP), landscape division index (DIVISION), contagion index (CONTAG), and percentage of like adjacencies (PLADJ; Table 2). Consequentially, the LUs positively related to PC1 were units diverse in composition, with a mixture in land cover classes, subdivided into fragments, mainly with urban and forest coverage. Principal component two (PC2), explained 22 % of the variation and was interpreted as a proxy for conservation (Figure 2). This component represents the urban gradient, due to the weight presented by the percentage of forest and urban coverage (Table 2).
Table 1 Landscape metrics per landscape units (LU) in San Cristóbal de Las Casas, Chiapas, México.
| Landscape metrics of composition | Landscape metrics of configuration | Distance (m) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LU name | ID | %URB | %FOR | %AGRO | %WET | SHDI | PR | NP | DIVISION | AI | CONTAG | PLADJ | DIS |
| Felipe Flores | FFL | 95.44 | 3.72 | 0.84 | 0 | 0.21 | 3 | 16 | 0.09 | 96.8 | 85.01 | 95.49 | 0 |
| C.31 de marzo | C31 | 78.84 | 9.77 | 11.39 | 0 | 0.66 | 3 | 27 | 0.37 | 92.12 | 56.04 | 90.59 | 1500 |
| San Nicolas | SNI | 60.01 | 13.67 | 28.91 | 0 | 0.93 | 3 | 17 | 0.6 | 90.84 | 41.93 | 89.17 | 1500 |
| Santa Cruz | SCR | 26.6 | 67 | 6.39 | 0 | 0.8 | 3 | 31 | 0.57 | 91 | 48.06 | 89.41 | 3500 |
| Cerrito SEDEM | CSE | 32.5 | 22.91 | 13.46 | 31.13 | 1.34 | 4 | 34 | 0.8 | 89.88 | 35.6 | 87.89 | 3500 |
| Reserva Moxviquil | MOX | 20.58 | 63.17 | 16.25 | 0 | 0.91 | 3 | 16 | 0.6 | 93.26 | 45.36 | 91.54 | 3500 |
| San Ramón | SRA | 65.26 | 5.83 | 28.91 | 0 | 0.8 | 3 | 18 | 0.55 | 92.35 | 49.63 | 90.73 | 3500 |
| San Felipe | SFE | 14.03 | 65.86 | 20.12 | 0 | 0.87 | 3 | 29 | 0.57 | 84.43 | 37.37 | 82.9 | 3500 |
| Albarrada Minas | ALM | 28.75 | 12.94 | 11.54 | 46.77 | 1.23 | 4 | 28 | 0.71 | 89.84 | 40.81 | 87.93 | 3500 |
| Alborada | ALB | 14.65 | 65.55 | 11.96 | 7.84 | 1.01 | 4 | 21 | 0.56 | 91.46 | 51.51 | 89.61 | 3500 |
| Parque Encuentro | ENC | 2.37 | 91.24 | 6.39 | 0 | 0.35 | 3 | 15 | 0.17 | 95.55 | 76.4 | 94.14 | 6000 |
| Cortijo | COR | 2.26 | 83.12 | 14.61 | 0 | 0.52 | 3 | 17 | 0.32 | 95.33 | 67.36 | 93.81 | 6000 |
| Parque Montetik | MON | 0 | 98.57 | 1.43 | 0 | 0.08 | 2 | 2 | 0.03 | 99.06 | 91.85 | 97.84 | 6000 |
| Periferia Huitepec | PHU | 12.69 | 63.75 | 23.56 | 0 | 0.89 | 3 | 41 | 0.59 | 83.44 | 36.38 | 81.91 | 6000 |
| Reserva Huitepec | RHU | 0 | 84.32 | 15.68 | 0 | 0.43 | 2 | 3 | 0.28 | 95.94 | 57.02 | 94.53 | 6000 |
Describing the landscape categories. The urban category was represented only by one unit (FFL; Figure 2). This category had the highest urban coverage and the highest aggregation index. The unit is located at the center of SCLC. In turn, the transition category was represented by heterogeneous units, with the highest richness of fragments (types of land cover). This category grouped units in process of urban transformation, with urban coverage between 28 % and 65 %; even though they retained proportions of other land cover classes; units are located between 1.5 and 3.5 km (Figure 2). The mosaic category gathered the units with the highest number of fragments; separated one from another, describing more than a continuum, a mosaic of forest, houses, agricultural plots and wetlands (Figure 2). Mosaic units were placed between 3.5 and 6 km from the city center. Conserved category was formed by units with the highest percentages of forest cover, and the lowest coverage of urban, and all are located at 6 km from the city center (Figure 2). A more detailed description of each category is presented in Supplementary material 3.
Rodent diversity, richness, and abundance. A sampling effort of 3,780 trap nights was carried out, during which 10 rodent species and 147 individuals were recorded (Supplementary material 4). Seven of these species, Oligoryzomys fulvescens, Peromyscus beatae, P. mexicanus, Reithrodontomys fulvescens, R. microdon, R. sumichrasti, and Sigmodon toltecus, belonged to the Cricetidae family and were considered native species; the remaining three species belong to the Muridae family and were considered non-native: Mus musculus, Rattus rattus and R. norvegicus. Regarding landscape categories, the most abundant species in the transition was S. toltecus and M. musculus was the most abundant in the mosaic (Figure 3). Rattus rattus was present throughout the study area, except in conserved units. In the conserved units, the most abundant species was P. mexicanus, which was also abundant in the mosaic units (Figure 3). The results of alpha diversity for each of the 15 LUs and for category of landscape are in Supplementary material 5.
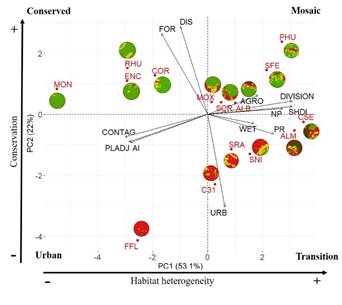
Figure 2 Plot loadings of PCA model, for landscape units by landscape metrics. Arrows = loadings per landscape metrics. Color poligons = landscape units.
Table 2 Scores of Principal Component Analysis (PCA) of landscape metrics and abundance of rodents in Landscapes Units (LU) in San Cristóbal de Las Casas, Chiapas, México.
| PCA Landscapes Unites (LU) | PCA Landscapes Unites (LU) and Rodent species | ||||
|---|---|---|---|---|---|
| Loadings | PC1 | PC2 | Loadings | PC1 | PC2 |
| URB | 0.193 | -0.935 | URB | 0.275 | -0.918 |
| FOR | -0.518 | 0.812 | FOR | -0.591 | 0.759 |
| AGRO | 0.562 | 0.191 | AGRO | 0.563 | 0.102 |
| WET | 0.516 | -0.09 | WET | 0.519 | 0.056 |
| PR | 0.738 | -0.195 | SHDI | 0.915 | 0.184 |
| NP | 0.835 | 0.061 | PR | 0.719 | -0.033 |
| DIVISION | 0.943 | 0.128 | NP | 0.833 | 0.182 |
| AI | -0.862 | -0.287 | DIVISION | 0.915 | 0.202 |
| CONTAG | -0.924 | -0.229 | AI | -0.846 | -0.325 |
| PLADJ | -0.881 | -0.28 | CONTAG | -0.895 | -0.27 |
| Eigenvalue | 6.379 | 2.639 | Eigenvalue | 7.713 | 3.782 |
| % of variance | 53.161 | 21.991 | % of variance | 35.06 | 17.191 |
| Cumulative % of variance | 53.161 | 75.152 | Cumulative % of variance | 35.06 | 52.251 |
According to the diversity of order 1 (1D), the units in the transition category have a diversity of 4.56 abundant species. In contrast to the 5.77 of the mosaic units (Supplementary material 5). This means that, among mosaic units, there are 1.21-fold more abundant species, in comparison to transition units, and 3-fold more than among conserved units (2.73). On the other hand, the highest-richness landscape categories were the transition (8 species) and the mosaic (7 species) units, while the lowest were the urban units (1 species; Figure 3). There are significant differences in richness between urban and transition categories (t = 4.2, P = 0.0005), and between urban and mosaic units (t = 3.28, P = 0.0041). Regarding abundance, coincidently, the most abundant categories are the transition ones (n = 76) and the mosaic (n = 43), and the lowest was the urban (n = 2). There are significant differences between abundance of urban and transition categories (t = 2.12, P = 0.0478), and between urban and mosaic (t = 2.99, P = 0.0077; Supplementary material 5).
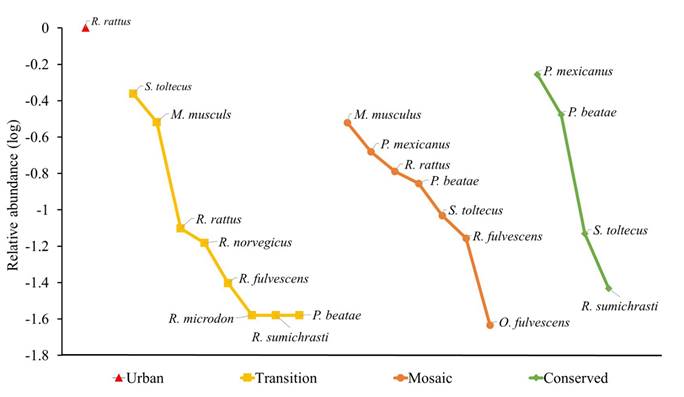
Figure 3 Range-abundance graphs by landscape categories in San Cristóbal de Las Casas, Chiapas, México.
The relationships between rodent species, landscape units, and the landscape metrics were also examined with an exploratory ordering analysis, via a PCA. The components 1 and 2 explained 35.1 % and 17.2 % of the variation, respectively (Figure 4). The presence of rodent species in SCLC is a function of two components: 1) habitat heterogeneity, defined by the landscape metrics: landscape division index (DIVISION), Shannon’s Diversity index (SHDI), contagion index (CONTAG), and aggregation index (AI; Table 2; Figure 4); and 2) urbanization, defined by the landscape metrics: forests cover (FOR), DIVISION, urban cover (URB), and aggregation index (AI; Table 2; Figure 4). There is a species’ grouping pattern that corresponds to the grouping pattern of landscape units (Figure 4). The units of mosaic category are associated to the presence of S. toltecus, R. fulvescens, and Oligoryzomys fulvescens. Urban units are characterized by the presence of Rattus rattus. Transition units group together due to sharing R. microdon, R. sumichrasti, R. norvegicus, and M. musculus. Finally, conserved units group due to P. beatae and P. mexicanus, although not exclusively (Figure 4).
Regarding beta diversity, the Sorensen index (βSOR) was 0.62; 52 % of this dissimilarity was due to species turnover (βSIM) and the remaining 48 %, due to the component resulting from nesting (βSNE). The landscape categories sharing the most species between them were the mosaic and transition units, with five species (P. beatae, R. fulvescens, S. toltecus, R. rattus, and Mus musculus). Conversely (according to the Sorensen dissimilarity index), the conserved LUs were the most dissimilar. Finally, six models explained the relation between metrics and rodent diversity. The landscape-composition variable that best explained species richness was the percentage of wetland coverage (AIC 55.9; Table 3). Abundance was better explained by the urban and forest cover variables (AIC 120.3) and Shannonʹs Diversity Index (SHDI, AIC 105.7; Table 3). In the case of landscape configuration metrics, DIVISION and CONTAG explained better the abundance of the species (AIC 98.094), and richness was better explained only by DIVISION (AIC 60.28).
Table 3 Estimated parameters for the best models of Generalized Linear Models for the effects of composition and configuration landscape on abundance and richness rodent in an urbanization gradient in San Cristóbal de Las Casas, Chiapas, México. SE = Standard error. AIC = Akaike criterion. P values < 0.05, are significant.
| Landscape composition | Parameters | Estimate | SE | AIC | Z-value | P-value |
|---|---|---|---|---|---|---|
| Abundance~URB+FOR | Intercept | 4.81 | 0.36 | 120.3 | 13.36 | < 0.00 |
| URB | -0.03 | 0 | -5.96 | 0 | ||
| FOR | -0.03 | 0 | -6.91 | 0 | ||
| Abundance~SHDI | Intercept | 0.64 | 0.27 | 105.7 | 2.37 | 0.01 |
| SHDI | 1.94 | 0.27 | 7.08 | 0 | ||
| q0 ~ WET | Intercept | 0.9 | 0.17 | 55.2 | 5.09 | 0 |
| WET | 0.01 | 0 | 1.85 | 0.06 | ||
| q0 ~ SHDI | Intercept | 0.09 | 0.44 | 51.9 | 0.2 | 0.83 |
| SHDI | 1.16 | 0.47 | 2.43 | 0.01 | ||
| Landscape configuration | ||||||
| Abundance ~ DIVISION + CONTAG | Intercept | -4.82 | 1.65 | 98 | -2.91 | 0 |
| DIVISION | 7.37 | 1.33 | 5.54 | 0 | ||
| CONTAG | 0.06 | 0.01 | 3.39 | 0 | ||
| q0~DIVISION | Intercept | 0.06 | 0.45 | 60.2 | 0.13 | 0.89 |
| DIVISION | 1.94 | 0.8 | 2.41 | 0.01 |
Discussion
In the middle-sized city of SCLC, the greatest diversity in terms of rodent species richness and abundance, was found in the categories of landscape mosaic and transition. In contrast, the lowest richness was in the urban and conserved categories. According to our results, there are more rodent species in the most heterogeneous landscapes, i. e., transition and mosaic landscape categories, where there is more than one type of land coverage and fragmentation occurs. The combination of urban, agricultural and wetland coverages contributed to the highest abundance and richness of rodent species in SCLC. This showed the importance of conserving those areas with wetland coverages and other natural vegetation coverages. Also, the majority of rodent species were distributed throughout the entire study area, regardless of the type of landscape, still some of them were more abundant in some landscape units.
The urbanization process in SCLC generates more than one landscape category, non-sequentially distributed, rather as spots in the study area and where the rodents are disseminated. The non-native rodents, M. musculus and R. rattus, were abundant in landscape categories with the highest proportions of urban coverage and were considered here as “urban dwellers”. This data matches reports in other cities, although of different sizes, in Argentina, United States and the Netherlands (Castillo et al. 2003; Gomez et al. 2009; Johnson et al. 2016; Maas et al. 2020). In Argentina, the community of non-native rodents was accompanied by native species, in smaller proportions (Castillo et al. 2003; Gomez et al. 2009). This structure of the rodent community was attributed to the urbanization processes of Latin American cities; which grow disorderly and settle in areas with natural vegetation. This drives native and non-native species to cohabit (Castillo et al. 2003), just like happens in San Cristóbal de Las Casas.
Some scholars attributed this community structure to the configuration of cities. For example, old cities (more than 100 years) that conserve riparian areas, small parks, or even large trees possess greater structural diversity, favoring a greater diversity of mammals (Parsons et al. 2018). Contrary to areas with less structural diversity or more homogeneous (e. g., monospecific forest areas) where there is less diversity of mammals. This reflects the patterns found in the city of SCLC, an old city preserving fragments with natural vegetation, agricultural plots and urban areas, generating a structurally complex landscapes, where native and non-native rodents cohabit.
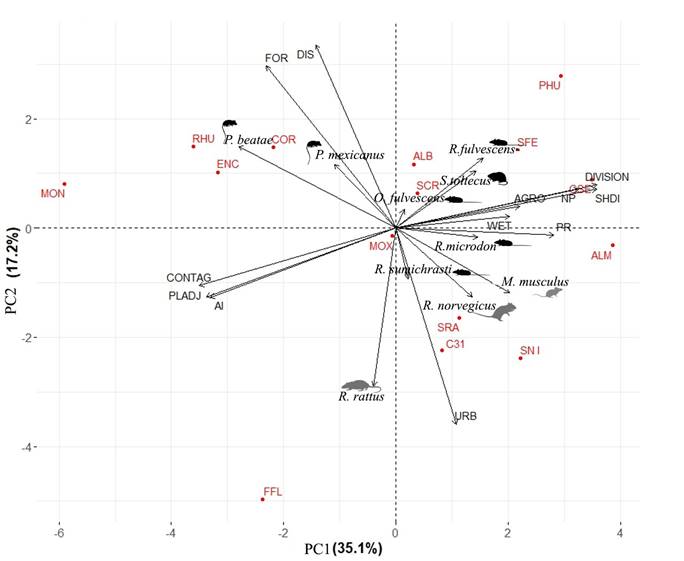
Figure 4 Plot loadings of PCA model. Arrows = loadings per landscape metrics and abundance of rodents. Points = landscape units. Black figures = native rodents. Gray figures = non native rodents.
Contrary to what was expected, the greatest richness and abundance of rodent species were concentrated in the mosaic and transition categories, and not in the conserved. This result coincides with that reported by García-Méndez et al. (2014) in SCLC. However, other studies carried out previously in the SCLC conserved units reported greater species richness. For example, Cruz Lara (2000), found five and three species of rodents in the Huitepec and Moxviquil reserves, respectively (versus one and two species reported in this study for the same reserves, respectively). Naranjo and Espinoza Medinilla (2001) reported a total of 11 species of the Cricetidae family and two of the Muridae, for the Huitepec Reserve. Finally, in 2018, Sántiz-Vázquez (2018)reported six species of cricetids in the Huitepec Reserve, two of which were also reported in this work. It is worth mentioning that these previous reports correspond to collections carried out systematically, monthly, during periods of six months or one year, which makes it likely that their numbers of species were greater than those reported in this study. Additionally, we believe that dogs, cats, chickens and other poultry (observed chasing mice) are contributing to the decrease in the number of rodents present in the reserve and park of SCLC.
Despite not being the richest sites, the conserved units harbored the greatest abundance of P. mexicanus and P. beatae, and no individuals of the non-native species M. musculus or Rattus species were found; although they had already been reported in the Huitepec Reserve (Naranjo and Espinoza Medinilla 2001). Even though the Mexican rodent, P. mexicanus, was present in two landscape categories in SCLC, these were the categories with the greatest forest coverage and with agricultural plots. This rodent species is widely distributed in the Mexican southeast and is considered a habitat generalist (Trujano-Ávarez and Álvarez-Castañeda 2010).
Our study indicates that habitat heterogeneity generates a landscape with intermediate disturbance, favoring an increase in species richness, as previously reported in other countries (Pardini et al. 2010). It is suggested that a structurally complex matrix in composition (i. e., heterogeneous landscapes) can increase rodent species richness within such landscapes. This argument was also suggested by San José et al. (2014); who reported that species richness may benefit from the matrix that generates fragmentation of the tropical forest as a source of food and shelter. Similarly, Medellín and Equihua (1998) showed that rodents “residing in the forest” may be able to use the corridors that form the matrix and benefit from the resources of places such as pastures or crops.
Given this study shows that rodent species (native and non-native) are abundant in human settlements, it is important to consider some sanitary measures; since R. norvegicus. R. rattus, and M. musculus are reservoirs of pathogens with zoonotic potential (Panti-May et al. 2012, 2018). However, native species such as P. mexicanus, S. toltecus, R. sumichrasti, and R. fulvescens are also reservoirs of zoonotic viruses potentially transmissible to humans and possible causes of hemorrhagic fever outbreaks (Inizan et al. 2010; Milholland et al. 2018). This is particularly important if we consider that S. toltecus and R. fulvescens are considered here as “urban visitors” and were found in houses, i. e., the contact with people is frequent.
This study highlights the necessity for more long-term studies focused on understanding landscape changes of the city of SCLC, as well as structural changes of the rodent community and their interactions with human populations. We suggest carrying out future monitoring activities that include virological analyses of the collected rodent specimens, to know the prevalence of zoonotic pathogens and establish the possible health risks of disorderly urban growth in SCLC.

