











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El monitoreo de la estructura poblacional, la consanguinidad, el tamaño efectivo de población y la probabilidad de origen de los genes, permite prevenir pérdidas de diversidad genética en las poblaciones bovinas1, particularmente en poblaciones bajo procesos de selección2. Por lo anterior, su estudio es necesario para implementar de manera más integral los programas de selección3.
En varios países se ha generado información acerca de la diversidad genética de sus poblaciones animales, tanto para razas locales como para transfronterizas (presentes en varios países), lo que ha sido ampliamente estudiado mediante análisis de pedigrí y usado en poblaciones de animales domésticos1-5. Sin embargo, en México, los estudios publicados utilizando el análisis de pedigrí en sus poblaciones han sido escasos6,7, por lo que el análisis de la estructura y variabilidad genética de sus poblaciones de animales domésticos debe ser una acción prioritaria de investigación en mejoramiento genético animal8.
Los métodos de selección comúnmente usados, como los basados en las predicciones de los valores genéticos obtenidos mediante el modelo animal, pueden incrementar los niveles de consanguinidad y disminuir la variabilidad genética de las poblaciones9. En México, las evaluaciones genéticas nacionales en razas de bovinos para carne iniciaron en 2001. En 2010 se reportó que 21 razas ya tenían evaluaciones genéticas10, en las cuales es importante conocer la diversidad genética presente. Por lo anterior, el objetivo de este estudio fue evaluar la diversidad genética de siete poblaciones de bovinos para carne en México, mediante el análisis de sus pedigríes.
Material y métodos
La información genealógica utilizada fueron las bases de datos de las poblaciones nacionales de bovinos Angus (AN), Brangus Negro (BN), Brangus Rojo (BR), Hereford (HE), Limousin (LI), Salers (SA) y Suizo Europeo (SE); provenientes de las respectivas asociaciones mexicanas de criadores de ganado de registro. El libro genealógico de cada raza incluyó animales nacidos en los periodos 1973-2017, 1964-2016, 1973-2017, 1935-2016, 1953-2017, 1976-2016 y 1927-2016 para AN, BN, BR, HE, LI, SA y SE, respectivamente. Los pedigríes originales se editaron para evitar la presencia de incongruencias, como registros duplicados, edades de los hijos mayores que las de sus progenitores o diferencias de edad no razonables biológicamente, y registros de animales como padres y como madres.
Los análisis de los pedigríes se llevaron a cabo mediante el programa Endog v4.8, donde se describen los detalles de los procedimientos usados para estimar los parámetros poblacionales estudiados11. Los análisis se realizaron para el total de los animales (PT) en cada raza, y para sus poblaciones de referencia (PR) se consideraron los animales en los 10 años más recientes (potencialmente vivos) de la población correspondiente. El significado de los parámetros poblacionales se describe a continuación.
Integridad del pedigrí
Los principales factores que influyen en la exactitud de los parámetros de las poblaciones usando el análisis de pedigrí son la integridad y nivel de completitud del pedigrí12. En este estudio, la integridad de los pedigríes se determinó mediante las estimaciones del número de generaciones completas (GC), número máximo de generaciones trazadas (MGT) y número de generaciones completas equivalentes (GE). Se consideró una generación completa cuando se conocían los 2 n antepasados de un individuo, mientras que las generaciones máximas trazadas relacionaron el número máximo de éstas que separan a un individuo de su ancestro más lejano11. Para un individuo dado, el número de generaciones completas equivalentes se obtuvo mediante la suma para todos los ancestros conocidos de los términos calculados como la suma de (1/2)n de todos los antepasados conocidos, donde n es el número de generaciones que separan al individuo de cada ancestro conocido13. Adicionalmente, como medida complementaria a la de integridad de los pedigríes, se calcularon los porcentajes de ancestros conocidos hasta la tercera generación parental.
Intervalo generacional (IG)
El IG fue definido como la edad promedio de los padres cuando nace su descendencia14. Este parámetro se calculó para las cuatro rutas gaméticas de selección: padre-hijo, padre-hija, madre-hijo y madre-hija.
Coeficientes de consanguinidad (F) y de relación genética aditiva promedio (CRP)
El F se calculó usando el algoritmo establecido por Meuwissen y Luo15, basado en el principio de que los fundadores no están emparentados y, además, en que su F es igual a cero. El CRP fue definido como la probabilidad de que un alelo escogido al azar de entre toda la población en el árbol genealógico pertenezca a un animal determinado11; por tanto, es una medida del porcentaje de genes que, en promedio, comparte cada individuo con el resto de los individuos del pedigrí. El cálculo del CRP se obtuvo del promedio de los coeficientes en la fila correspondiente al individuo en la matriz de relaciones genéticas aditivas “A”14,16.
Tamaño efectivo de población (N e ) y tasa de incremento en consanguinidad (ΔF)
El N
e
es el número de individuos que darían lugar a la tasa calculada de consanguinidad, si fueran criados bajo el sistema de la población idealizada17. El N
e
representa las relaciones de parentesco entre los números de machos y hembras que contribuyen genéticamente en la población y el parámetro es inversamente proporcional a la ΔF5. El incremento en consanguinidad se estimó mediante:
Probabilidad de origen de los genes
El pedigrí de un individuo puede ser rastreado para calcular la contribución a su genoma de su eventual fundador o ancestro. Un fundador fue definido como un ancestro con progenitores desconocidos, pero cuando sólo se conoce un progenitor, el progenitor desconocido es considerado como fundador. La preservación de la diversidad genética de los fundadores hacia la población de estudio puede ser medida por sus contribuciones12.
Se consideró como número efectivo de fundadores (f e ) al número de animales fundadores que contribuyeron por igual para producir la diversidad genética observada en la población20. Si todos los fundadores contribuyen por igual a la descendencia, f e es igual al número de fundadores (f); por el contrario, si la contribución es desigual, f e es de menor valor12,20. Por otro lado, se consideró como número efectivo de ancestros (f a ) al número mínimo de ancestros (fundadores o no) necesarios para explicar la diversidad genética completa de una población12. Este parámetro toma en cuenta la posible existencia de cuellos de botella en el pedigrí, por el uso extensivo de algunos individuos en ciertas generaciones. Finalmente, se consideró el parámetro número efectivo de genomas fundadores equivalentes (f g ) al número de fundadores que podría esperarse produjeran la misma diversidad genética de la población bajo estudio, si los fundadores estuvieran igualmente representados y no hubiera ocurrido pérdida de alelos21. Adicionalmente, considerando las contribuciones genéticas marginales12, se obtuvieron los números de ancestros que explican el 50 % de los genes, y la contribución del principal ancestro.
Resultados y discusión
Censos en las poblaciones
En el Cuadro 1 se muestra una descripción general de los animales considerados en cada raza estudiada. El número de animales utilizados por raza en el total del pedigrí fue variable (aproximadamente entre 13,000 y 185,000 para las PT, y entre 4,000 y 37,000 para las PR). Los promedios de hijos por semental en cada población fluctuaron entre 6 y 31, pero en las PR los valores fueron prácticamente el doble que en las respectivas PT, excepto en la raza SE que fueron la mitad. Los promedios de hijos por vaca oscilaron entre 1.8 y 2.7. Los porcentajes de animales con padre o madre desconocidos en las PT fue similar en todas las poblaciones, y en promedio fue alrededor de 16 %, oscilando para las diferentes poblaciones entre 8 (BN) y 33 % (HE); los valores disminuyeron sustancialmente para los animales nacidos en los 10 años más recientes (PR).
Cuadro 1 Descripción general del número (N) de animales (A) en los pedigríes de las siete poblaciones bovinas mexicanas estudiadas, para la población total y la población de referencia* (dentro de paréntesis)
| Concepto | Población total (población de referencia) | ||||||
|---|---|---|---|---|---|---|---|
| AN | BN | BR | HE | LI | SA | SE | |
| NA total | 73,271 | 68,474 | 12,925 | 13,248 | 53,221 | 14,065 | 184,788 |
| (34,433) | (30,356) | (6,329) | (4,444) | (22,804) | (4,885) | (37,245) | |
| N de sementales | 8,224 | 4,408 | 1,842 | 1,408 | 4,220 | 1,382 | 5,157 |
| (905) | (696) | (280) | (123) | (385) | (67) | (481) | |
| N de madres | 30,948 | 25,090 | 6,124 | 4,587 | 20,587 | 5,377 | 59,609 |
| (8,357) | (5,735) | (1,895) | (890) | (4,067) | (511) | (5,042) | |
| NA con padre desconocido | 10,539 | 5,612 | 2,133 | 4,413) | 6,219 | 1,768 | 21,845 |
| (988) | (204) | (5) | (168) | (213) | (7) | (3) | |
| NA con madre desconocida | 11,420 | 5,048 | 2,129 | 4,450 | 6,812 | 1,765 | 24,337 |
| (1,282) | (333) | (9) | (164) | (389) | (5) | (4) | |
| N de hijos por semental (NHS) | 7.63 | 14.26 | 5.86) | 6.27 | 11.14 | 8.90 | 31.60 |
| (16.67) | (17.72) | (10.90) | (14.40) | (20.87) | (17.91) | (16.36) | |
| Máximo NHS | 412 | 561 | 312 | 150 | 710 | 358 | 1,620 |
| (246) | (293) | (219) | (71) | (182) | (65) | (155) | |
| N de hijos por vaca | 2.00 | 2.53 | 1.76 | 1.92 | 2.25 | 2.29 | 2.70 |
| (2.00) | (1.97) | (1.80) | (1.92) | (2.14) | (1.97) | (1.78) | |
| N de hatos | 202 | 152 | 86 | 48 | 214 | 118 | 648 |
| (175) | (122) | (70) | (37) | (132) | (21) | (308) | |
AN= Angus; BN= Brangus Negro; BR= Brangus Rojo; HE= Hereford; LI= Limousin; SA= Salers; SE= Suizo Europeo.
* Población de referencia (10 años más recientes).
Integridad de los pedigríes
Los valores promedio de los GC, MGT y GE para las poblaciones bovinas estudiadas se muestran en el Cuadro 2. De acuerdo con los GE obtenidos para la PT, las poblaciones con mayor información sobre las genealogías fueron BN y SE (GE > 4), mientras que en las demás poblaciones, los valores estimados (2.0 a 3.9) fueron inferiores a los obtenidos en otras investigaciones con razas europeas transfronterizas para carne (4.0 a 9.3)22,23. Los valores para los MGT y GC en las razas estudiadas tuvieron un patrón similar al observado para los GE. Con excepción de HE, los MGT estimados para las demás poblaciones fueron superiores o similares (5.4 a 10.3) a los estimados (4.2 a 6.9)24,25) en razas bovinas criollas (Costeño con Cuernos, Blanco Orejinegro, Romosinuano y Sanmartinero) y transfronterizas (Guzerat). Sin embargo, inferiores (12 a 14)22) a los publicados para otras razas bovinas transfronterizas (Chianina, Marchigniana y Romagnola). Los GC estimados en las poblaciones (2.4 a 3.0) fueron mayores que los obtenidos en bovinos Guzerat (1.7)25, con excepción de la población HE, donde se obtuvo un valor menor (1.5). Los GE, GC y MGT obtenidos en la PR fueron mayores que los de las respectivas PT y comparables con los publicados en otras poblaciones de bovinos22-25.
Cuadro 2 Números de generaciones máximas trazadas (MGT), completas (GC) y completas equivalentes (GE) conocidas en los pedigríes de las siete poblaciones bovinas mexicanas estudiadas, para la población total y la población de referencia* (dentro de paréntesis)
| Indicador | Población total (población de referencia) | ||||||
|---|---|---|---|---|---|---|---|
| AN | BN | BR | HE | LI | SA | SE | |
| MGT | 8.09 (11.61) | 10.28 (13.48) | 5.40 (8.09) | 3.32 (6.22) | 7.65 (11.16) | 6.83 (10.61) | 8.50 (13.03) |
| GC | 2.52 (3.41) | 3.05 (3.66) | 2.29 (3.25) | 1.45 (2.55) | 2.41 (3.50) | 2.58 (3.93) | 2.37 (3.64) |
| GE | 3.87 (5.32) | 5.89 (7.51) | 3.38 (4.91) | 2.03 (3.55) | 3.76 (5.42) | 3.84 (5.62) | 4.28 (6.38) |
AN= Angus; BN= Brangus Negro; BR= Brangus Rojo; HE= Hereford; LI= Limousin; SA= Salers; SE= Suizo Europeo. * Población de referencia (10 años más recientes).
* Población de referencia (10 años más recientes).
Según Maignel et al26, el criterio más apropiado para caracterizar la información del pedigrí es el GE, mientras que los MGT y GE son indicadores complementarios que dan mejor idea del grado de profundidad del pedigrí. Las proporciones de ancestros conocidos sobre todas las generaciones trazadas (GE) obtenidas en este estudio para las PT, indican que algunas estimaciones de parámetros poblacionales pudieran tener limitaciones al compararse con las de otras poblaciones de otros estudios. Sin embargo, con excepción de HE, para las PR de las demás poblaciones estos valores fueron mayores (4.9 a 7.5), lo que muestra los esfuerzos recientes de los criadores en el registro de información genealógica de sus animales.
El grado de integridad de los pedigríes en las poblaciones estudiadas se confirmó con las estimaciones de porcentajes de ancestros conocidos (Cuadro 3). La PT de HE presentó los menores porcentajes de progenitores conocidos hasta la tercera generación (38 a 67 %); mientras que las demás poblaciones presentaron porcentajes razonables de ancestros conocidos en la primera (>84 %), segunda (>71 %) y tercera (>61 %) generación parental. En las PR, los porcentajes de progenitores conocidos hasta la tercera generación aumentaron sustancialmente (>90 %), con excepción de HE, donde los valores fueron entre 77 y 90 %. Los porcentajes de ancestros conocidos publicados en otros estudios han sido variables; valores superiores al 90 % de ancestros conocidos en la tercera generación parental han sido reportados para la población Marchigiana27, mientras que en las poblaciones españolas Bruna de los Pirineos y Asturiana de los Valles se han estimado valores cercanos al 3 %14. Por otro lado, las poblaciones de este estudio mostraron un patrón similar de valores entre las rutas paterna y materna, lo que sugiere que la captura de registros fue equitativa entre sexos. Conocer la profundidad del pedigrí es importante, ya que esto influye en la estimación de parámetros que son sensibles a la completitud de la información genealógica disponible12; por lo anterior, para HE se esperarían subestimaciones en algunos de los parámetros analizados, principalmente en los F.
Cuadro 3 Porcentajes de ancestros conocidos de las siete poblaciones bovinas mexicanas estudiadas, para la población total y la población de referencia* (dentro de paréntesis)
| Ancestros ¥ | Población total (población de referencia) | ||||||
|---|---|---|---|---|---|---|---|
| AN | BN | BR | HE | LI | SA | SE | |
| Progenitores | |||||||
| S | 86 (98) | 92 (100) | 84 (100) | 67 (98) | 88 (99) | 87 (100) | 88 (100) |
| M | 84 (97) | 93 (100) | 84 (99) | 66 (98) | 87 (99) | 87 (100) | 87 (100) |
| Abuelos | |||||||
| SS | 80 (98) | 90 (99) | 76 (99) | 56 (97) | 84 (99) | 82 (100) | 84 (100) |
| MS | 80 (96) | 89 (99) | 75 (97) | 56 (97) | 84 (99) | 80 (100) | 82 (100) |
| SM | 74 (95) | 86 (97) | 71 (98) | 49 (88) | 77 (97) | 78 (100) | 78 (100) |
| MM | 73 (93) | 87 (98) | 71 (98) | 49 (88) | 76 (96) | 78 (100) | 77 (100) |
| Bisabuelos | |||||||
| SSS | 75 (96) | 87 (99) | 69 (99) | 46 (90) | 81 (99) | 78 (100) | 82 (100) |
| MSS | 76 (97) | 86 (97) | 69 (97) | 46 (90) | 81 (97) | 72 (100) | 77 (98) |
| SMS | 72 (96) | 84 (96) | 65 (96) | 44 (91) | 76 (99) | 71 (94) | 77 (98) |
| MMS | 73 (96) | 84 (96) | 66 (96) | 44 (90) | 76 (89) | 72 (98) | 74 (99) |
| SSM | 69 (94) | 84 (97) | 65 (97) | 40 (82) | 73 (97) | 74 (100) | 74 (99) |
| MSM | 69 (95) | 82 (95) | 65 (96) | 39 (82) | 72 (96) | 74 (99) | 72 (100) |
| SMM | 64 (92) | 81 (96) | 61 (96) | 38 (77) | 64 (95) | 70 (98) | 66 (99) |
| MMM | 63 (90) | 82 (96) | 61 (96) | 38 (77) | 64 (88) | 71 (100) | 63 (98) |
AN= Angus; BN= Brangus Negro; BR= Brangus Rojo; HE= Hereford; LI= Limousin; SA= Salers; SE= Suizo Europeo. * Población de referencia (10 años más recientes). ¥ S= semental; M= madre.
Intervalo generacional
Los IG estimados para las poblaciones estudiadas en las cuatro rutas de selección posibles se muestran en el Cuadro 4. En promedio, para las PT los IG de las razas estudiadas fluctuaron entre 5.1 y 6.5 años, incrementándose sus valores en todas las PR (5.5 a 7.2 años). Las pérdidas de diversidad genética se producen a mayor velocidad con menores IG; sin embargo, provocan mayor progreso genético por unidad de tiempo16. En general, los IG promedios de las poblaciones de este estudio fueron menores a los de otros estudios25,28,29 con bovinos para carne de razas transfronterizas (Guzerat, Charolais, Limousin, Hereford, Angus y Simmental) y locales (Alentejana). Sin embargo, para LI y SE en la PT, y para las PR, los IG fueron mayores en la ruta de padres para producir hijos que en las demás rutas gaméticas, lo que sugiere el uso de sementales por periodos prolongados, con posiblemente un uso de pocos reproductores e inseminación artificial. Este patrón de comportamiento con mayores IG para la ruta padre-hijo fue similar al publicado en Angus28 y Suizo Pardo30. En investigaciones con razas bovinas locales, algunos estudios14,24 han estimado IG en las rutas paternas menores que los de las maternas, atribuyéndolo al reemplazo temprano de los sementales.
Cuadro 4 Intervalos generacionales (años) estimados para las cuatro rutas gaméticas de selección en animales reproductores de las siete poblaciones bovinas mexicanas estudiadas, para la población total y la población de referencia* (dentro de paréntesis)
| Población total (población de referencia) | |||||||
| Ruta gamética | AN | BN | BR | HE | LI | SA | SE |
| Padre-hijo | 6.19 (6.64) | 5.25 (5.82) | 5.54 (6.50) | 4.93 (6.71) | 7.67 (11.12) | 5.09 (7.57) | 8.19 (8.90) |
| Padre-hija | 5.41(5.71) | 5.50 (5.71) | 5.56 (5.57) | 5.36 (5.59) | 7.45 (8.23) | 4.98 (6.92) | 5.79 (6.99) |
| Madre-hijo | 5.19 (5.55) | 5.35 (5.53) | 5.26 (5.51) | 5.31 (5.85) | 5.91 (6.09) | 5.04 (6.86) | 6.95 (6.38) |
| Madre-hija | 5.27 (5.59) | 5.74 (5.82) | 5.14 (5.21) | 5.45 (5.54) | 5.50 (5.63) | 5.19 (6.71) | 5.95 (6.40) |
| Promedio | 5.4 (5.7) | 5.6 (5.8) | 5.3 (5.5) | 5.4 (5.7) | 6.5 (7.2) | 5.1 (6.9) | 6.4 (6.7) |
AN= Angus; BN= Brangus Negro; BR= Brangus Rojo; HE= Hereford; LI= Limousin; SA= Salers; SE= Suizo Europeo. * Población de referencia (10 años más recientes).
Pirchner31 menciona que los IG en las rutas padre-hijo= 6.5 a 9.0, padre-hija= 4.5 a 6.5, madre-hijo= 5.0 a 6.5 y madre-hija= 5.0 a 6.5 años, son valores que pueden considerarse como estándar en bovinos para carne. De acuerdo con esa clasificación, en general, los valores estimados en este estudio fluctuaron en esos rangos (con excepción de la PR de LI para las rutas padre-hijo y padre-hija, donde se obtuvieron IG mayores a los estándares); sin embargo, para BN, BR, HE y SA en la ruta padre-hijo fueron inferiores a los estándares (<5.6 años) en la PT, lo que sugiere recambios cortos y continuos de los sementales activos.
Consanguinidad y relación genética aditiva promedio
Los F y los CRP en las poblaciones estudiadas se muestran en el Cuadro 5. Los valores promedio de los F para las PT de las poblaciones estudiadas fueron relativamente bajos (0.9 a 3.5 %), por lo que podría esperarse poca manifestación de la depresión endogámica; sin embargo, en los años recientes algunas poblaciones muestran mayores niveles de F (>4 %; BN y BR), posiblemente debido al uso más intensivo de pocos sementales. Resultados intermedios a los valores de F obtenidos en este estudio han sido publicados para poblaciones de bovinos Tropicarne (1.8 %) y Suizo Europeo (1.5 %) de México6, y en varias poblaciones pequeñas de bovinos (Alistana, Morucha, Pirenaica, Blanco Orejinegro, Romosinuano, Marchigniana, Bonsmara, Charolais, Limousin y Simmental, entre otras) reportadas por diversos autores14,24,27,32. Los aumentos importantes en el porcentaje de animales consanguíneos y su consanguinidad promedio en la PR con relación a la PT, para las razas BN y SA, sugieren poca atención en la asignación de apareamientos de estas poblaciones en los años recientes.
Cuadro 5 Porcentajes de animales consanguíneos (PAF), del coeficiente de relación genética aditiva promedio (CRP), y del promedio de coeficientes de consanguinidad en la población (F-PO) y en los animales consanguíneos (F-AC) de las siete poblaciones bovinas mexicanas estudiadas, para la población total y la población de referencia* (dentro de paréntesis)
| Población total (población de referencia) | |||||||
|---|---|---|---|---|---|---|---|
| AN | BN | BR | HE | LI | SA | SE | |
| PAF | 47.4 (75.8) | 77.2 (94.9) | 38.5 (60.4) | 13.7 (30.9) | 29.3 (44.9) | 39.2 (71.4) | 56.3 (95.7) |
| F-PO | 0.9 (1.4) | 3.5 (4.8) | 2.6 (4.0) | 0.9 (1.8) | 0.9 (0.8) | 0.9 (2.2) | 1.5 (2.4) |
| F-AC | 2.0 (1.9) | 4.6 (5.0) | 6.8 (6.6) | 6.2 (5.7) | 2.9 (0.8) | 2.3 (3.1) | 2.7 (2.5) |
| CRP | 0.5 (0.7) | 5.2 (6.5) | 1.7 (2.7) | 0.3 (0.5) | 0.5 (1.1) | 1.0 (1.5) | 1.1 (1.4) |
AN= Angus; BN= Brangus Negro; BR= Brangus Rojo; HE= Hereford; LI= Limousin; SA= Salers; SE= Suizo Europeo. * Población de referencia (10 años más recientes).
Considerando los valores de todos los parámetros asociados con los F, las poblaciones BN y HE pudieran presentar problemas futuros, esta última debido a posibles subestimaciones en los parámetros, ya que tiene las mayores proporciones de padres y madres desconocidos (Cuadro 1), y menores integridades de sus pedigríes (Cuadros 2 y 3).
Con la excepción de BN y BR, los CRP de las demás poblaciones fueron bajos (<1.5 %), lo que sugiere que la consanguinidad podría decrecer en las poblaciones si se practica migración entre hatos14 dentro de poblaciones. Los CRP estimados en BN y BR (>1.7 %) sugiere problemas futuros para evitar apareamientos entre individuos no emparentados en esas poblaciones. Los CRP son indicadores de la consanguinidad a largo plazo en una población; cuando estos alcanzan valores mayores que los F, los apareamientos deben ser cuidadosamente planificados, para evitar apareamientos entre animales emparentados33 y sugieren problemas de la población para mantener la variabilidad genética34, como es el caso de BN del presente estudio. Al respecto, Gutiérrez et al14 mencionan que un CRP bajo junto con un F alto en poblaciones, indica que la proporción de apareamientos dentro de hatos es alta y que el intercambio de animales entre hatos es poco frecuente. Una alternativa sugerida35 es que los reproductores con un menor valor individual del CRP deben preferirse como progenitores de las próximas generaciones, para minimizar los incrementos de los F de los animales en el largo plazo.
La selección de animales basada en el mejor predictor linear insesgado (BLUP) también favorece la selección de animales emparentados y, por tanto, da lugar a valores altos de F y CRP36. En todas las poblaciones del presente estudio, los porcentajes de animales consanguíneos y los F a través del tiempo mostraron una tendencia gradual creciente (Figura 1); además, la implementación de evaluaciones genéticas en las poblaciones estudiadas, acicatea la necesidad de establecer estrategias para el control de la consanguinidad. En los 10 años más recientes, aunque en forma fluctuante para algunas razas, la tendencia fue de mantener los F, con la excepción de SA donde continuó el incremento gradual de F. Lo anterior sugiere atención de algunos criadores en la asignación de los apareamientos en los años recientes.
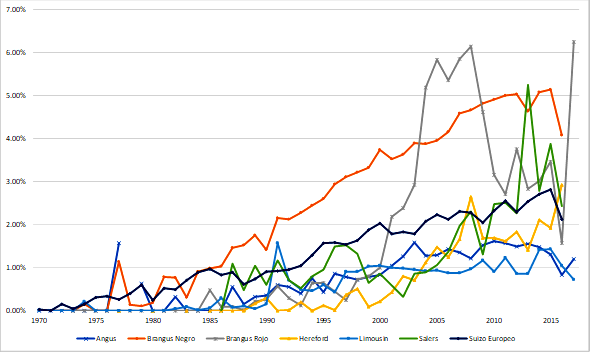
Tasa de incremento en consanguinidad y tamaño efectivo de la población
La ΔF y el N e estimados en las razas de bovinos mexicanos estudiadas se muestran en el Cuadro 6. El N e osciló entre 24 (SA) y 192 (LI). La ΔF se comportó de forma análoga al N e , debido a que son inversamente proporcionales. Según la FAO37, para la conservación y mejora de las poblaciones es deseable contar con un N e mínimo de 50, que es equivalente al 1 % en la ΔF. De las poblaciones del presente estudio, sólo dos poblaciones (BR y SA) se encontraron por debajo del nivel recomendado por la FAO. Henson38 señaló que la heterocigosidad o diversidad genética presente en cada generación decrece a una tasa acelerada en poblaciones con N e menor que 100 y que el incremento en homocigosis como consecuencia de un N e reducido resulta en la pérdida de habilidad adaptativa, depresión endogámica y por último la extinción. Esta situación sería preocupante en cuatro de las razas analizadas en este estudio (BN, BR, HE y SA); sin embargo, dado el carácter transfronterizo de las razas estudiadas, es posible la migración y flujo de germoplasma entre países. Bajo este escenario, la entrada de genes nuevos a las poblaciones de bovinos de México, podría traer beneficios a la diversidad genética nacional; sin embargo, de no hacerse, podría afectar negativamente en el caso de diseminación masiva de material genético de pocos reproductores por diversas técnicas reproductivas en animales altamente seleccionados.
Cuadro 6 Tasas de incremento en consanguinidad promedio
| Población (raza) | |||||||
|---|---|---|---|---|---|---|---|
| Parámetro | AN | BN | BR | HE | LI | SA | SE |
|
|
0.32 | 0.74 | 1.04 | 0.65 | 0.26 | 2.08 | 0.45 |
| N e | 155 | 68 | 48 | 77 | 192 | 24 | 110 |
AN= Angus; BN= Brangus Negro; BR= Brangus Rojo; HE= Hereford; LI= Limousin; SA= Salers; SE= Suizo Europeo
El N e estimado a partir del incremento en consanguinidad individual ha sido evaluado en pedigríes simulados y reales de distintas especies39,40,41. Danchin-Burge et al40 realizaron una comparación entre tres distintas metodologías utilizadas en la obtención del N e para ocho razas de ganado lechero francés, una mediante la obtención de la ΔF a partir de la regresión lineal del F sobre el tiempo, otra mediante el cálculo del incremento individual en el F, y la otra basada en el incremento en la coancestría. En general, el método de incremento individual en el F presentó valores mayores que los otros dos, donde la menor diferencia fue estimada en las poblaciones más grandes y con mejor calidad de pedigrí. Según Cervantes et al39, la ΔF individual depende en gran parte de la calidad de pedigrí y necesita al menos cinco generaciones completas equivalentes para hacerse constante. Al respecto, se comenta que cuando la información de pedigrí es incompleta, puede provocar sesgos al estimar la ΔF y, en consecuencia, del N e 12.
Danchin-Burge et al40 evaluaron el N e a partir del ΔF individual, obteniendo valores de N e entre 67 y 243 en ocho razas francesas de bovinos lecheros, mientras que en otro trabajo41 obtuvieron un estimado de 94 en la raza Gir de Brasil. Por otro lado, valores reportados de N e por otros métodos han sido extremadamente variables, donde las razas locales y lecheras especializadas son las que presentan los menores valores14,23,26,28,42. En general, las poblaciones analizadas en el presente estudio se encontraron dentro del intervalo de la mayoría de valores publicados, excepto SA que tuvo un N e inferior.
Probabilidad de origen del gen
Los parámetros descriptivos de la probabilidad de origen del gen de las siete poblaciones estudiadas se muestran en el Cuadro 7. En general, las estimaciones de los parámetros relacionados con la probabilidad de origen de los genes indican que la presencia de contribuciones desbalanceadas del uso de los animales fundadores, los cuellos de botella genéticos y la deriva genética, han tenido un efecto importante en las poblaciones actuales (10 años más recientes), con la consecuente pérdida de diversidad genética.
Cuadro 7 Parámetros descriptivos de la probabilidad de origen de los genes estimados en las siete poblaciones bovinas mexicanas estudiadas, para la población total y la población de referencia* (dentro de paréntesis)
| Población total (población de referencia) | |||||||
|---|---|---|---|---|---|---|---|
| Parámetro ¥ | AN | BN | BR | HE | LI | SA | SE |
| f | 10,168 (9,019) | 4,827 (3,622) | 2,131 (1,855) | 2,439 (1,618) | 5,862 (4,214) | 1,728 (1,134) | 11,886 (3,412) |
| f e | 541 (431) | 113 (97) | 192 (125) | 413 (308) | 519 (299) | 331 (237) | 307 (246) |
| f a | 224 (168) | 33 (27) | 55 (32) | 254 (159) | 199 (96) | 105 (62) | 166 (87) |
| f a / f e , % | 41.4 (39.0) | 29.2 (27.8) | 28.6 (25.6) | 61.5 (51.6) | 38.3 (32.1) | 31.7 (26.2) | 54.1 (35.4) |
| f g | 189 | 19 | 54 | 361 | 187 | 90 | 93 |
| C max , % | 3.2 (3.4) | 13.0 (14.6) | 9.3 (11.9) | 1.5 (2.3) | 2.6 (4.2) | 4.4 (6.3) | 3.1 (3.8) |
| N 50 | 177 (102) | 16 (12) | 44 (16) | 94 (56) | 100 (190) | 44 (25) | 42 (31) |
AN= Angus; BN= Brangus Negro; BR= Brangus Rojo; HE= Hereford; LI= Limousin; SA= Salers; SE= Suizo Europeo. * Población de referencia (10 años más recientes).
¥ f= número total de fundadores; f e = número efectivo de fundadores; f a = número efectivo de ancestros; C max = contribución del principal ancestro; N 50 = número de ancestros que contribuyen con el 50 % de los genes a la población; f g = número efectivo de genomas fundadores.
Los valores de los f fueron mayores que los f e , lo que indica disminución en diversidad genética, debido a desbalances en el uso de los animales fundadores. En porcentaje, la contribución desigual de los fundadores (f e /f) en las PT fue más importante en AN, BN y SE (2.3 a 5.3 %) que en BR, HE, LI y SA (8.9 a 19.2 %); valores que fueron similares en las respectivas PR, con sólo ligera mejora detectada para SE. La contribución desigual de los fundadores es indicador de posibles incrementos en consanguinidad para generaciones futuras, lo que reflejaría la pérdida de diversidad genética (reducción de heterocigosidad y pocas variantes alélicas por locus) presente en los fundadores20. En el presente estudio, la contribución muy desbalanceada de los fundadores implica que un reducido número de animales se están utilizando como reproductores en BR, LI y SA, mientras que en ese indicador para HE, adicionalmente pudiera tener influencia la poca integridad del pedigrí. Al respecto, Boichard et al12 señalan que en poblaciones con baja calidad de pedigrí puede existir una gran cantidad de fundadores fantasmas, lo que provoca sobreestimaciones del f y f e . Algunas poblaciones de razas de bovinos para carne transfronterizas (Chianina, Marchigniana, Romagnola y Nelore) han mostrado una baja relación (0.23 a 1.09 %) en los parámetros mencionados22,43. No obstante, valores similares a los de este estudio han sido reportados en otros estudios con razas bovinas para carne (Charolais, Limousin, Hereford, Angus y Simmental)23,28.
En el presente estudio, el f a estimado en las siete poblaciones bovinas analizadas fue menor que su respectivo f e , lo que indica disminución en la diversidad genética por la presencia de cuellos de botella genéticos en todas las poblaciones, definidas a partir de su fundación. Los efectos del cuello de botella fueron más importantes en los pedigríes de las PT para AN, BN, BR, LI y SA (f a /f e <50%), mientras que para HE y SE se estimaron valores mayores que 50 %. Lo anterior podría sugerir un efecto pequeño de los cuellos de botella genéticos en esas últimas poblaciones; sin embargo, la información debe considerarse con cautela en HE, dado que esa población es la que presentó peor integridad de pedigrí. De acuerdo con algunos autores12, los parámetros derivados de la probabilidad de origen del gen son sensibles a la calidad del pedigrí, aunque en menor grado que el F, siendo el f a el más afectado. Los cuellos de botella fueron más estrechos (f a /f e entre 26 y 52%) para todas las poblaciones en los 10 años más recientes (PR), lo que indica la necesidad de establecer medidas para incrementar la diversidad genética.
A partir de las contribuciones marginales consideradas en la estimación de los f a , es posible identificar a los ancestros más sobresalientes por su contribución marginal relativa a la diversidad genética de la población en estudio12. Con base en ello, el número de ancestros que explicaron el 50 % de la diversidad genética presente para la PT en cada una de las siete poblaciones estudiadas fue variable, fluctuando desde 16 individuos en BN hasta 177 en AN (Cuadro 7), y con disminuciones para las respectivas PR. Las altas contribuciones marginales de algunos ancestros provocaron que el número de individuos que explican el 50 % de la diversidad genética presente en las poblaciones bajo estudio muestren una relativamente reducida base genética. Diversos autores han reportado cantidades menores que 10 individuos6,32,40,42, valores menores que en las poblaciones estudiadas en la presente investigación. Sin embargo, estas contribuciones de los ancestros a los genes de las respectivas poblaciones deben ser consideradas por los criadores, debido a que la alta presencia de genes de unos pocos reproductores en las poblaciones podría llevar a un incremento de apareamientos entre animales emparentados, consecuentemente a la generación de individuos consanguíneos y pérdida de variabilidad genética.
En general, los f a fueron mayores que los f g de las PT, sugiriendo disminuciones, de ligeras a moderadas, en la diversidad genética por la presencia de pérdidas aleatorias de alelos en todas las poblaciones (deriva genética), desde los progenitores fundadores a sus progenies, por lo que se recomienda el uso de un rango más amplio de sementales.
Conclusiones e implicaciones
La diversidad genética presente en las poblaciones de bovinos mexicanas evaluadas fue variable. Aunque en todas las poblaciones los coeficientes de consanguinidad promedio fueron relativamente bajos, en algunas se detectaron problemas de pocos reproductores utilizados, tamaño efectivo de población pequeño y potenciales problemas con apareamientos entre individuos emparentados, presentando (en mayor o menor grado) pérdidas en diversidad genética por contribuciones desbalanceadas del uso de los animales fundadores, cuellos de botella genéticos y deriva genética. Considerando los parámetros estudiados, las poblaciones AN, LI y SE fueron las que presentaron la mayor diversidad genética. En SA se requiere incrementar su tamaño efectivo de población, el uso de mayor diversidad en reproductores y la atención en la asignación de apareamientos. Las razas BN, BR y HE presentaron las mayores pérdidas de diversidad genética e incrementos en consanguinidad, por lo que se deben implementar estrategias para mantener su variabilidad genética futura; específicamente en HE se debe mejorar la integridad y completitud de su pedigrí, ya que afecta la estimación de algunos de los parámetros y su interpretación apropiada. A pesar que se detectan esfuerzos recientes de los criadores en el registro de información genealógica de sus animales, se recomienda continuar con el monitoreo de parámetros relacionados con la diversidad genética y la captura de información de pedigrí. Además, aunque se trata de razas transfronterizas, es necesario implementar estrategias para incrementar sus tamaños efectivos, evitar los incrementos en consanguinidad y las pérdidas de diversidad genética, mediante los diseños de apareamientos y el uso estratégico de migración con genes provenientes de otros países.

