











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Cedrela odorata L. es la segunda una especie de importancia forestal en México; al ser considerada madera preciosa, posee gran aceptación social por parte de productores para el establecimiento de plantaciones forestales (Sánchez-Monsalvo et al., 2003) y el precio de la madera es atractivo (Ramírez-García et al., 2008). Al igual que otros países, México utiliza principalmente especies exóticas para el establecimiento de plantaciones comerciales ya que se genera poca información sobre el manejo de las especies nativas (dos Santos et al., 2022).
Pese a su importancia, las plantaciones comerciales de Cedrela odorata se ven limitadas debido al ataque del barrenador de yemas Hypsipyla grandella Zeller, que es la principal plaga de las Meliáceas americanas (Howard y Merida, 2004; Grignola et al., 2021). La larva se alimenta de los brotes tiernos y ocasiona el rompimiento de la yema apical provocando que el árbol se bifurque, lo que puede reducir su crecimiento afectando su valor comercial (Cibrián Tovar, 1995).
Como parte de la búsqueda de mecanismos de resistencia al ataque del barrenador, el Instituto Nacional de Investigaciones Forestales y Agropecuarias (Inifap) inició la recolección de germoplasma de Cedrela odorata para el establecimiento ensayos de progenies en la costa del Golfo y sureste de México con el propósito de obtener plantas mejoradas de esta especie con alta productividad y resistentes al daño de H. grandella. Posteriormente se llevó a cabo el establecimiento de ensayos clonales propagados por injerto para evaluar la estimación de parámetros genéticos cuantitativos en caracteres de crecimiento.
Debido al tipo de daño que ocasiona el barrenador en el fuste, se infiere que un árbol alto con fuste recto presentó tolerancia al ataque del barrenador en su estado juvenil, por tal motivo al evaluar el crecimiento y la rectitud del fuste se determina tolerancia de un genotipo a ataques del insecto (Briceño-Vergara, 1997). Por otro lado, para registrar y proteger la propiedad intelectual del material vegetal producto del mejoramiento se deben determinar descriptores morfológicos que tengan repetibilidad entre sitios de evaluación y por tanto alto control genético (Bennadji, 2003).
La estimación de parámetros genéticos en ensayos clonales es primordial para definir estrategias de mejoramiento y selección de genotipos superiores (White et al., 2007). Los parámetros genéticos como la heredabilidad y correlaciones genéticas determinan el control genético bajo el cual se encuentran las características de interés (Ipinza, 1998). La heredabilidad estima en qué medida un carácter es transmisible a la progenie y permite inferir su respuesta a la selección (White et al., 2007). Las correlaciones genéticas son una medida del grado de enlace genético entre dos o más caracteres en el mismo individuo, su causa principal es la pleiotropía, que es la propiedad que tiene un gen de afectar a dos o más caracteres (Falconer y Mackay, 2017).
Cuando las características se encuentran bajo fuerte control genético responden a la selección. Pero no es adecuado seleccionar un genotipo probado en un solo sitio y utilizarlo en condiciones ambientales distintas ya que puede existir un comportamiento diferencial en las plantaciones, esto es interacción genotipo-ambiente (White et al., 2007). Los genotipos propagados deben ser estables en diferentes ambientes, lo que se evalúa a través de determinar la magnitud de la interacción genotipo-ambiente de una característica entre sitios donde se evalúan los mismos genotipos (Johnson, 1997).
Objetivos
El objetivo del presente estudio fue estimar heredabilidad, correlación genética y fenotípica e interacción genotipo-ambiente de 14 caracteres morfológicos y de crecimiento de Cedrela odorata L. en dos ensayos clonales a siete años. Estos parámetros genéticos son esenciales para establecer programas que mejoren la resistencia al barrenador y el crecimiento de las plantas.
Materiales y métodos
Descripción del área de estudio
En 2012, se establecieron dos ensayos clonales en ambientes diferentes. El primero se localiza en terrenos de la empresa “Agropecuaria Santa Genoveva” en San Francisco, Campeche (Sitio 1) en 19º 33’ 26.53” N, 90º 01’ 33.96” O (Fig. 1), a 82 m s.n.m., con clima Aw0 (Instituto Nacional de Estadística y Geografía [Inegi], 2008), el más seco de los cálidos subhúmedos con lluvias en verano (García, 2004), precipitación de 1300 mm anuales y temperatura media anual de 23 °C. El tipo de suelo es Rendzina (International Union of Soil Sciences [IUSS], 2014), con textura arcillosa y de origen calcáreo (Honorato-Salazar y Sánchez-Monsalvo, 2019). El uso de suelo es bosque cultivado (Inegi, 2018). El segundo ensayo se estableció en propiedad privada en Isla, Veracruz (Sitio 2) en 18° 04’ 50.21” N, 95° 32’ 0.75” O (Fig. 1), a 56 m s.n.m., con clima Aw1 (w) (INEGI, 2008), intermedio de los cálidos subhúmedos (García, 2004), precipitación de 2000 mm anuales y temperatura media anual de 25 °C, con lluvias en verano. El tipo de suelo es Andosol vítrico (IUSS, 2014), ácido y con textura arenosa (Honorato-Salazar y Sánchez-Monsalvo, 2019). El uso de suelo es agricultura de temporal semipermanente (Inegi, 2018).
Figura 1 Mapa de ubicación de dos ensayos clonales de Cedrela odorata L. en Santa Genoveva, Campeche e Isla, Veracruz.
Diseño y establecimiento de los ensayos clonales El material vegetal usado para el establecimiento de los dos ensayos clonales fueron yemas para la propagación por injertos y fue obtenido de genotipos sobresalientes en caracteres de crecimiento de ensayos de procedencias-progenies establecidos en El Palmar, Veracruz; San Felipe Bacalar, Quintana Roo y El Tormento, Campeche. El ensayo 1, de Santa Genoveva, Campeche, se estableció con riego por goteo y el control de malezas se hizo con herbicida biológico-orgánico. En el ensayo dos en Isla, Veracruz, se realizó preparación mecanizada del terreno y se aplicó riego durante el primer mes de establecimiento de la plantación. Los dos ensayos clonales fueron establecidos en un diseño experimental de bloques completos al azar con 90 clones y seis repeticiones, con parcelas de un solo árbol (Pedersen et al., 1993).
Variables evaluadas
Se midió altura total (ALT) con un estadal y diámetro normal (DIAM, a 1.3 m de altura del fuste) con cinta diamétrica. Se estimó el volumen del fuste con la ecuación de Sánchez y García (2009):
La rectitud del fuste se evaluó considerando su inclinación, esto es su desviación respecto a la línea vertical que debería seguir. Se usó la aplicación Spirit level (Keuwlsoft, 2020) de un celular colocado a 1.5 m de altura del árbol; la observación se expresó en grados. Esto permite trabajar con una variable continua, al obtener el valor en grados, y evitar el uso de categorías en la inclinación del fuste. Se evaluaron ocho caracteres cualitativos y dos cuantitativos (Méndez-Espinoza y Sánchez-Monsalvo, 2012), para ello se recolectaron cuatro hojas completas de la parte media de la copa, una por cada punto cardinal del árbol y se promediaron los datos. Los caracteres cuantitativos fueron: largo de la hoja (cm), ancho de la hoja (cm) y longitud del peciolo (cm) con cinta graduada; longitud del peciolulo (mm), longitud de los foliolos (mm), ancho de los foliolos secundarios (mm) con vernier; número de pares de foliolos secundarios e índice de forma del foliolo secundario (relación entre largo/ancho). Como caracteres cualitativos se evaluaron la forma del fuste a la altura de 1.30 m y el tipo de ramificación. La forma del fuste se categorizó en: circular, deltoide, elipsoide, ovada, pentagonal, romboide y semicircular; el tipo de ramificación se clasificó en dicotómica, monopódica, policotómica y simpódica.
Análisis estadístico
Se realizó un análisis por sitios en conjunto con el procedimiento MIXED del paquete estadístico SAS (SAS Institute Inc., 2002). Se usó el algoritmo de máxima verosimilitud restringida para la estimación de varianza (Littell et al., 2016). El modelo estadístico usado fue:
donde:
Y ijkl = valor de la observación del l-ésimo clon de la j-ésima procedencia en el k-ésimo bloque (repetición) en el i-ésimo sitio
μ= media poblacional
Si = efecto fijo del i-ésimo sitio
B j(i) = efecto fijo del bloque anidado en el i-ésimo sitio
P k = efecto fijo de la k-ésima procedencia
SP ik = efecto fijo de la interacción entre el i-ésimo sitio con la k-ésima procedencia
BP ijk =efecto fijo de la interacción del j-ésimo bloque anidado en el i-ésimo sitio con la k-ésima procedencia
C l(k) = efecto aleatorio del l-ésimo clon anidado en la k-ésima procedencia
SC ikl = efecto aleatorio de la interacción del i-ésimo sitio y el l-ésimo clon anidado en la k-ésima procedencia
ε ijkl = error asociado a dichos efectos
Además, se realizó el análisis por sitio debido a las diferencias entre los ensayos. Se usó un modelo similar al análisis en conjunto, sin los efectos de los sitios y sus interacciones:
Estimación de parámetros genéticos
Con los valores de varianza se estimó la heredabilidad en sentido amplio para rametos (
donde:
b = media armónica del número de bloques (individuos evaluados): 4.09 y 5.61 para los sitios 1 y 2, respectivamente.
Se calcularon correlaciones fenotípicas mediante la correlación de Pearson, a través del procedimiento CORR de SAS (SAS Institute Inc., 2002), y las correlaciones genéticas (r gxy ) calculadas a partir de las fórmulas descritas por Falconer y Mackay (2017):
donde:
donde:
Para evaluar el desempeño de los clones en ambos sitios se estimó la interacción genotipo-ambiente. Con el fin de remover los efectos de escala se estandarizaron los datos mediante el procedimiento STANDARD de SAS (Hodge et al., 1996). Una vez hecho esto, se calcularon la varianza deσ 2clones (σ2 c ) y la varianza de la interacción sitio x clon (σ2 sc ). con el procedimiento MIXED de SAS y se estimaron correlaciones genéticas tipo B como medida de la interacción genotipo-ambiente (Yamada, 1962):
Se calculó la contribución relativa de cada clon a la interacción genotipo-ambiente mediante el método de ecovalencia (Wricke, 1962) con el fin de conocer los genotipos más inestables:
donde:
W i = ecovalencia del i-ésimo clon
Y ij = posición jerárquica del i-ésimo clon en el j-ésimo sitio
Y i. = media del i-ésimo clon en el j-ésimo sitio
Y .j = media del j-ésimo sitio
Y.. = media general
El volumen fue utilizado para determinar el valor de ecovalencia y la contribución a la interacción genotipo-ambiente. Se evaluó el efecto de la exclusión de los clones más interactivos sobre la estimación de los rparámetros heredabilidad y correlaciones genéticas tipo B (r B ).
Resultados y discusión
Se encontraron diferencias significativas entre sitios (p < 0.01) para 11 de las 14 variables evaluadas: diámetro, altura, volumen, inclinación, forma del fuste, longitud y ancho de hoja, longitud de peciolo y de peciolulo, longitud y ancho de foliolos secundarios. En el sitio 1 (Santa Genoveva, Campeche) los valores medios de la mayoría de las características fueron superiores a los obtenidos en el sitio 2 (Isla, Veracruz), a excepción de la inclinación, ancho de hoja, longitud del peciolo y del peciolulo (Tabla 1). Los valores mayores en Santa Genoveva se pueden atribuir a que durante los primeros cinco años de la plantación se aplicó riego por goteo; así con mayor disponibilidad de agua se obtiene más crecimiento (Huber y Trecaman, 2004). Asimismo, con la mayor mortalidad en Santa Genoveva (supervivencia 79% vs. 94%), los individuos dispusieron de menor competencia para desarrollarse (Sánchez-Vargas et al., 2004).
Tabla 1 Heredabilidad a nivel individual (Hi2), a nivel clonal (Hc2) y coeficiente de variación genética (CVG) por sitio en dos ensayos clonales de Cedrela odorata L. de siete años en Santa Genoveva, Campeche y en Isla, Veracruz.
| Santa Genoveva, Campeche (Sitio 1) | Isla, Veracruz (Sitio 2) | |||||||||
| Variable | Promedio | Intervalo |
|
|
CV G | Promedio | Intervalo |
|
|
CV G |
| DIAM* | 16.0 | 6.7- 26.3 | 0.09 | 0.28 | 6.6 | 11.0 | 3.3-18.4 | 0.09 | 0.35 | 9.6 |
| ALT* | 8.5 | 4.0-12.0 | 0.05 | 0.19 | 1.3 | 6.3 | 3.0- 10.0 | 0.15 | 0.49 | 2.5 |
| VOL* | 110.0 | 10.0-321.0 | 0.07 | 0.24 | 203.0 | 40.0 | 1.9-137.0 | 0.20 | 0.58 | 220.0 |
| INC* | 6.5 | 0.5- 30.4 | 0.10 | 0.31 | 63.0 | 9.7 | 0.5-38.5 | 0.06 | 0.27 | 87.0 |
| FFUS* | 4.4 | 1.0- 7.0 | 0.07 | 0.23 | 9.2 | 4.5 | 1.0-7.0 | 0.03 | 0.16 | 4.5 |
| TRAM | 2.2 | 1.0- 4.0 | 0.02 | 0.06 | 0.6 | 2.2 | 1.0-4.0 | 0.03 | 0.16 | 0.9 |
| LHOJA* | 35.3 | 21.5-51.1 | 0.09 | 0.29 | 7.1 | 31.1 | 18.9-51.8 | 0.24 | 0.64 | 15.5 |
| AHOJA* | 16.5 | 11.5-22.6 | 0.07 | 0.23 | 1.5 | 16.9 | 10.6-23.5 | 0.24 | 0.64 | 4.8 |
| LPEC* | 6.1 | 3.2- 9.8 | 0.11 | 0.34 | 2.5 | 6.4 | 3.8-9.5 | 0.22 | 0.61 | 3.4 |
| LPU* | 8.3 | 2.7-18.4 | 0.17 | 0.46 | 14.2 | 7.5 | 2.5-16.1 | 0.51 | 0.86 | 31.6 |
| NPFS | 8.1 | 5.0-12.0 | 0.09 | 0.30 | 1.3 | 8.4 | 5.8-12.5 | 0.13 | 0.46 | 1.6 |
| LFS* | 88.0 | 58.0-116.0 | 0.09 | 0.29 | 11.8 | 82.0 | 52.0-158.0 | 0.25 | 0.65 | 31.1 |
| AFS* | 31.7 | 20.1-57.7 | 0.15 | 0.41 | 9.6 | 30.9 | 17.2-46.3 | 0.41 | 0.80 | 24.8 |
| IFFS | 2.8 | 1.5- 3.8 | 0.11 | 0.34 | 0.5 | 2.7 | 1.7-4.0 | 0.35 | 0.75 | 1.6 |
DIAM = Diámetro (cm), ALT = Altura (m), VOL = Volumen (dm3), INC = Inclinación, FFUS = Forma del fuste, TRAM = Tipo de ramificación LHOJA y AHOJA = Longitud (cm) y ancho (cm) de hoja, LPEC y LPU = Longitud de peciolo (cm) y de pecioluloσ2(mm), ̅ NPFS = Número de pares de foliolos secundarios, LFS, AOFS y IFFS = Longitud (mm), ancho (mm) e índice de forma de foliolo secundario (largo/ancho). CVG = (/ )100. * Diferencias significativas (p < 0.01).
Los árboles de Isla presentaron mayor inclinación. No se tiene la certeza de atribuir esta respuesta al ataque de H. grandella, dado que no se realizó una evaluación de ello en la etapa de mayor afectación que son los primeros tres años (Navarro et al., 2004). Sin embargo, puede inferirse que hubo mayor daño por el barrenador antes de la evaluación actual, ya que este insecto ocasiona deformación del fuste (Cornelius y Watt, 2003). Asimismo, en el segundo sitio hay mayor precipitación, lo que genera más cantidad de brotes tiernos, esto es mayor disponibilidad de alimento para la larva del barrenador, lo que puede propiciar un mayor daño.
Heredabilidad por sitio individual y clonal
El control genético de las variables evaluadas fue de bajo a moderado. En Santa Genoveva la heredabilidad estimada para rametosH 2 en diámetro, altura y volumen tuvieron valores de i de H0.09,2 0.05 y 0.07 y para heredabilidad media de clones c 0.28, 0.19 y 0.24. Los valores de heredabilidad fueron más bajos comparados los registrados por Navarro et al. (2004) en un ensayo de 115 familias de 21 procedenciash asociado con café en Costa Rica, quienes señalan i2 dentro de las procedencias de 0.12 y 0.20 para diámetro y altura respectivamente, Igualmente fueron más bajos que los registrados por Sampayo-Maldonado et al. (2019), en un ensayo clonal de dos años generado por a que en dicha evaluación hubo mayor número genotipos evaluados. La razón por la que dicho experimento presentó mayor heredabilidad puede atribuirse a que cuando se tiene mayor número de repeticiones aumenta el tamaño de muestra, lo que incrementa la precisión de la estimación o se disminuye la variabilidad microambiental, asimismo cuando no hay 100% de supervivencia, la competencia entre las plantas no es la misma (Falconer y Mackay 2017).
Asimismo, los valores de
El tipo de ramificación presentó el valor más bajo de
La longitud del peciolulo presentó los valores más altos con
En Isla, 11 de 14 valores de heredabilidad para clones fue mayor que en Santa Genoveva. En heredabilidad de rametos del diámetro, altura y volumen fueron
El control genético de la inclinación del presente estudio fue de
El volumen, la inclinación, la longitud del peciolulo y la longitud de foliolos secundarios presentaron coeficientes de variación genética mayores a 10% en los dos sitios, además la longitud de hoja y el ancho de foliolos secundarios también presentaron coeficientes mayores a 10% en Isla, lo que implica considerable variabilidad en esas característicasCVen ambos sitios. En esta investigación los valores de G son mayores que los registrados por Hernández-Máximo et al. (2016) en un ensayoCV de progenies de la misma especie. Un mayor valor de Gestá asociado con mayor potencial de ganancia genética (Houle, 1992) y es de esperar mayor progreso en el mejoramiento genético dado por la mayor respuesta a la selección (Falconer y Mackay, 2017).
Correlaciones genéticas y fenotípicas
En los dos sitios las correlaciones genéticas y fenotípicas obtenidas entre diámetro, altura y volumen fueron altas y positivas. El sitio 1 presentó valores de 0.76 a 0.90 en correlación fenotípica y 0.93 a 0.97 en correlación genética. En el sitio 2 las correlaciones fenotípicas variaron de 0.74 a 0.97 y las genéticas fueron de 0.74 a 1.00 (Tablas 2 y 3). Esto indica que las variables están asociadas entre sí y que al seleccionar por altura o diámetro se obtendrá una respuesta correlacionada alta y positiva en volumen. Estos datos coinciden con los registrados por Hernández-Máximo et al. (2016), quienes a la edad de 11 años hallaron correlaciones genéticas en variables de crecimiento de 0.81 a 0.99. Igualmente coinciden con Sánchez Monsalvor et al. (2003), que a los cinco años registraron valores de gde 0.91 a 0.95, ambos estudios llevados a cabo en la Cedrela odorata en Tezonapa, Veracruz.
Tabla 2 Correlaciones genéticas (debajo de la diagonal) y fenotípicas (sobre la diagonal) de variables evaluadas en un ensayo clonal de Cedrela odorata L. a los siete años en Santa Genoveva, Campeche (Sitio 1).
| DIAM | ALT | VOL | INC | FFUS | TRAM | LHOJA | AHOJA | LPEC | LPU | NPFS | LFS | AFS | IFFS | |
| DIAM | 0.76 | 0.96 | -0.14 | -0.27 | -0.21 | 0.09 | -0.03 | 0.07 | 0.10 | -0.10 | 0.02 | 0.09 | -0.09 | |
| ALTURA | 0.91 (0.03) | 0.84 | -0.17 | -0.23 | 0.02 | 0.15 | -0.08 | 0.07 | 0.06 | -0.11 | 0.04 | 0.09 | -0.06 | |
| VOL | 0.97 (0.01) | 0.93 (0.02) | -0.16 | -0.23 | -0.15 | 0.10 | -0.05 | 0.08 | 0.11 | -0.12 | 0.01 | 0.09 | -0.1 | |
| INC | -0.44 (0.11) | -0.35 (0.14) | -0.71 (0.07) | -0.01 | -0.03 | 0.02 | -0.02 | -0.05 | -0.01 | -0.04 | -0.03 | 0.10 | -0.14 | |
| FFUS | -0.34 (0.14) | -0.65 (0.11) | -0.33 (0.15) | -0.28 (0.14) | 0.07 | -0.04 | 0.04 | -0.05 | 0.04 | 0.06 | 0.03 | -0.02 | 0.04 | |
| TRAM | -1.00 (0.03) | -0.72 (0.16) | -0.93 (0.04) | -0.18 (0.25) | -0.97 (0.02) | -0.08 | -0.07 | -0.04 | -0.04 | -0.02 | -0.02 | -0.07 | 0.06 | |
| LHOJA | -0.07 (0.14) | 0.52 (0.12) | -0.04 (0.15) | 0.59 (0.09) | -0.29 (0.14) | 0.59 (0.17) | 0.61 | 0.54 | 0.31 | 0.31 | 0.56 | 0.36 | 0.13 | |
| AHOJA | 0.00 (0.16) | 0.44 (0.15) | -0.19 (0.16) | -0.10 (0.15) | -0.36 (0.15) | -0.39 (0.25) | 0.72 (0.07) | 0.36 | 0.29 | 0.21 | 0.73 | 0.48 | 0.15 | |
| LPEC | -0.03 (0.13) | 0.26 (0.15) | 0.17 (0.13) | -0.26 (0.11) | -0.17 (0.14) | 0.18 (0.24) | 0.41 (0.11) | -0.17 (0.12) | 0.25 | -0.14 | 0.24 | 0.23 | -0.01 | |
| LPU | 0.45 (0.09) | 0.20 (0.13) | 0.48 (0.09) | -0.75 (0.05) | 0.27 (0.11) | -1.00 (0.00) | -0.33 (0.10) | -0.09 (0.14) | 0.32 (0.09) | -0.10 | 0.24 | 0.33 | -0.16 | |
| NPFS | 0.02 (0.14) | 0.25 (0.16) | -0.36 (0.13) | 0.66 (0.07) | -0.04 (0.15) | N.E. | 0.13 (0.13) | 0.00 (0.12) | -0.30 (0.14) | -0.95 (0.01) | 0.12 | -0.20 | 0.33 | |
| LFS | 0.16 (0.14) | 0.44 (0.14) | 0.18 (0.15) | -0.34 (0.12) | -0.13 (0.15) | -0.33 (0.24) | 0.40 (0.11) | -0.11 (0.15) | -0.27 (0.12) | -0.28 (0.10) | -0.32 (0.12) | 0.57 | 0.29 | |
| AFS | 0.01 (0.12) | 0.03 (0.14) | -0.07 (0.13) | -0.03 (0.11) | -0.39 (0.11) | -0.71 (0.11) | 0.43 (0.09) | 0.84 (0.04) | 0.11 (0.11) | 0.35 (0.08) | -0.48 (0.09) | 0.52 (0.08) | -0.59 | |
| IFFS | 0.12 (0.13) | 0.50 (0.12) | 0.31 (0.13 | -0.38 (0.11) | 0.49 (0.11) | 0.89 (0.05) | -0.17 (0.12) | 1.02 (0.09) | -0.38 (0.10) | -0.63 (0.06) | 0.17 (0.12) | 0.21 (0.12) | -0.73 0.05 |
DIAM = Diámetro, ALT = Altura, VOL = Volumen, INC = Inclinación, FFUS = Forma del fuste, TRAM = Tipo de ramificación LHOJA y AHOJA = Longitud y ancho de hoja, LPEC y LPU = Longitud de peciolo y de peciolulo, NPFS = Número de pares de foliolos secundarios, LFS, AFS y IFFS = Longitud, ancho e índice de forma de foliolo secundario (largo/ancho). Error estándar de las correlaciones genéticas entre paréntesis. N.E. No estimable.
Tabla 3 Correlaciones genéticas (debajo de la diagonal, error estándar de las correlaciones genéticas) y fenotípicas (sobre la diagonal) de variables evaluadas en un ensayo clonal de Cedrela odorata L. a los siete años en Isla, Veracruz (Sitio 2).
| DIAM | ALT | VOL | INC | FFU | TRA | LHOJA | AHOJA | LPEC | LPU | NPFS | LFS | AFS | IFFS | |
| DIAM | 0.74 | 0.97 | -0.13 | -0.29 | -0.22 | 0.22 | 0.07 | 0.17 | 0.16 | 0.13 | 0.08 | 0.02 | 0.05 | |
| ALT | 0.88 (0.01) | 0.86 | -0.20 | -0.22 | -0.12 | 0.15 | 0.06 | 0.13 | 0.18 | 0.08 | 0.04 | 0.02 | 0.00 | |
| VOL | 1.00 (0.00) | 0.89 0.01 | -0.16 | -0.26 | -0.17 | 0.21 | 0.05 | 0.16 | 0.16 | 0.12 | 0.06 | 0.00 | 0.04 | |
| INC | -0.43 (0.06) | -0.76 0.04 | -0.52 (0.06) | -0.07 | -0.03 | 0.01 | 0.1 | 0.11 | 0.01 | -0.05 | 0.13 | 0.12 | -0.01 | |
| FFU | -0.46 (0.07) | -0.26 0.11 | -0.45 (0.08) | 0.39 (0.13) | 0.04 | -0.06 | -0.04 | -0.02 | -0.01 | -0.08 | -0.06 | -0.01 | -0.05 | |
| TRA | -0.56 (0.07) | -0.60 0.07 | -0.66 (0.06) | -0.07 (0.15) | 0.14 (0.19) | -0.07 | -0.07 | -0.12 | -0.09 | 0.03 | -0.03 | -0.08 | 0.06 | |
| LHOJA | 0.44 (0.04) | 0.14 0.06 | 0.41 (0.04) | 0.05 (0.07) | -0.43 (0.08) | -0.31 0.09 | 0.46 | 0.39 | 0.24 | 0.47 | 0.55 | 0.44 | 0.05 | |
| AHOJA | -0.02 (0.05) | -0.13 0.06 | -0.05 (0.05) | 0.04 (0.08) | -0.43 (0.08) | -0.23 0.09 | 0.61 (0.03) | 0.36 | 0.33 | 0.15 | 0.73 | 0.43 | 0.21 | |
| LPEC | 0.15 (0.05) | 0.04 (0.06) | 0.15 (0.05) | 0.65 (0.05) | -0.02 (0.09) | -0.17 0.10 | 0.49 (0.04) | 0.56 (0.03) | 0.32 | -0.06 | 0.34 | 0.30 | -0.01 | |
| LPU | 0.24 (0.03) | 0.49 (0.03) | 0.32 (0.03) | -0.15 (0.04) | 0.16 (0.09) | -0.26 0.05 | 0.26 (0.03) | 0.32 (0.03) | 0.20 (0.03) | -0.04 | 0.19 | 0.31 | -0.14 | |
| NPFS | 0.18 (0.06) | -0.29 (0.07) | 0.03 (0.06) | -0.17 (0.09) | -0.85 (0.03) | 0.14 0.12 | 0.27 (0.06) | -0.30 (0.05) | -0.33 (0.06) | -0.16 (0.04) | 0.17 | -0.03 | 0.19 | |
| LFS | -0.03 (0.05) | -0.19 (0.05) | -0.10 (0.05) | 0.20 (0.07) | 0.13 (0.09) | -0.38 0.08 | 0.57 (0.03) | 0.99 (0.00) | 0.53 (0.03) | 0.10 (0.03) | -0.26 (0.05) | 0.57 | 0.31 | |
| AFS | -0.05 (0.03) | 0.09 (0.04) | -0.01 (0.04) | 0.33 (0.05) | -0.24 (0.06) | -0.14 0.07 | 0.58 (0.02) | 0.62 (0.02) | 0.54 (0.03) | 0.44 (0.02) | -0.26 (0.04) | 0.53 (0.02) | -0.58 | |
| IFFS | 0.02 (0.04) | 0.09 (-0.23) | -0.07 (0.04) | -0.30 (0.06) | 0.41 (0.06) | -0.17 0.08 | -0.23 (0.04) | 0.06 (0.04) | -0.26 (0.04) | -0.42 (0.02) | 0.04 (0.05) | 0.159 (0.04 | -0.74 (0.01) |
DIAM = Diámetro, ALT = Altura, VOL = Volumen, INC = Inclinación, FFUS = Forma del fuste, TRAM = Tipo de ramificación LHOJA y AHOJA = Longitud y ancho de hoja, LPEC y LPU = Longitud de peciolo y de peciolulo, NPFS = Número de pares de foliolos secundarios, LFS, AFS y IFFS = Longitud, ancho e índice de forma de foliolo secundario (largo/ancho). Error estándar de las correlaciones genéticas entre paréntesis. según Falconer y Mackay (2017).
Sin embargo, es necesario tomar en cuenta la inclinación del fuste, ya que un árbol seleccionado deberá tener alto crecimiento e inclinación reducida. La variable inclinación del fuste presentó correlación negativa con variables de crecimiento en ambos sitios. Los individuos con mayor diámetro, altura y volumen tendrán menor inclinación. Aunque también es probable que los valores de inclinación observados estén asociados a la respuesta emitida ante el ataque del barrenador; lo anterior se infiere porque se sabe que el ataque induce la emisión de brotes laterales y como consecuencia el fuste se ramifica y deforma (Hilje y Cornelius, 2001).
Entre variables de crecimiento y morfológicas las correlaciones fueron muy variables, tanto positivas de moderadas a bajas, como negativas y bajas. Las correlaciones genéticas entre longitud del peciolulo con diámetro, altura y volumen en Santa Genoveva fueron de 0.45, 0.20 y 0.48, respectivamente; y las de Isla de 0.24, 0.49 y 0.32, consideradas como moderadas. Esto indica que la longitud del peciolulo puede servir para la selección de individuos con características de crecimiento superiores, al menos en una primera etapa de selección de genotipos, antes de las pruebas de campo para evaluar el crecimiento.
En Santa Genoveva, la correlación genética entre índice de forma del foliolo secundario fue moderada y positiva con altura y volumen (0.50 y 0.61). Un mayor índice denota foliolos más delgados y alargados, lo que indicaría que se podrían obtener individuos de mayor altura y volumen al propagar progenies de árboles con foliolos secundarios más delgados y alargados. Sin embargo, en Isla las correlaciones fueron casi nulas (0.02, 0.09 y -0.07).
La correlación genética entre número de pares de foliolos secundarios y ancho del foliolo secundario resultó alta y negativa con valor de -0.95 en Santa Genoveva, a diferencia de Isla que fue de -0.16. Aunque la mayoría de las correlaciones fenotípicas y genéticas presentaron diferencias entre los dos sitios, varias fueron consistentes en ambos, tal es el caso de las variables de crecimiento, longitud de hoja con longitud del foliolo secundario, longitud del peciolo con índice de forma del foliolo secundario, longitud del foliolo secundario con ancho de foliolo secundario e índice de forma del foliolo secundario y ancho del foliolo secundario.
Las diferencias en correlaciones genéticas entre sitios se han encontrado en Pinus patula (Morales González et al., 2013). Esto se explica como "respuesta correlacionada asimétrica", la que se puede deber a un cambio en las frecuencias genéticas de esas variables entre ambos ensayos (Bohren et al., 1966).
Los valores de correlaciones genéticas tipo B (rB) variaron de 0.29 hasta 1.00 (Tabla 4). De acuerdo con Robertson (1959), un valor de B> 0.8 indica reducida interacción genotipo-ambiente y un comportamiento estable en la característica de interés entre ambos sitios de evaluación. Las variables evaluadas que presentaron una rB por encima de 0.80 fueron inclinación y forma del fuste, tipo de ramificación, longitud del peciolulo, número de pares de foliolos secundarios y longitud y ancho del foliolo secundario. Las características que presentaron bajar interacción genotipo-ambiente con valores aceptables de B (0.74 a 0.79) fueron diámetro, altura, volumen, índice de forma del foliolo secundario, ya que valores cercanos a 1 indican que los genotipos superiores lo son igualmente en otros ambientes (White et al., 2007). Esto indica que para estos caracteres es posible seleccionar genotipos superiores en caracteres de crecimiento y estables en ambos sitios de evaluación. Las variables con valores más bajos de rB fueron longitud de hoja (0.64), ancho de hoja (0.53) y longitud del peciolo (0.29). Los valores encontrados en variables de crecimiento son menores a los obtenidos en los mismos experimentos en un análisis realizado a un año, con datos de rB de 0.90 en altura y 0.85 en diámetro (Salazar-García et al., 2015).
Tabla 4 Correlaciones genéticas Tipo B (rB) y componentes de varianza en dos ensayos clonales de Cedrela odorata L. de siete años ubicados en Santa Genoveva, Campeche e Isla, Veracruz.
| Característica | Correlaciones genéticas tipo B | Componentes de varianza | |
| σ 2 c | σ 2 s*c | ||
| Diámetro | 0.74 | 0.096 | 0.033 |
| Altura | 0.74 | 0.055 | 0.019 |
| Volumen | 0.79 | 0.080 | 0.022 |
| Inclinación del fuste | 0.97 | 0.065 | 0.002 |
| Forma del fuste | 1.00 | 0.060 | 0.000 |
| Tipo de ramificación | 1.00 | 0.031 | 0.000 |
| Longitud de hoja | 0.64 | 0.080 | 0.045 |
| Ancho de hoja | 0.53 | 0.059 | 0.052 |
| Longitud de peciolo | 0.29 | 0.037 | 0.089 |
| Longitud de peciolulo | 0.90 | 0.249 | 0.028 |
| No. pares de foliolos secundarios | 0.94 | 0.080 | 0.005 |
| Longitud foliolo secundario | 0.38 | 0.055 | 0.087 |
| Ancho foliolo secundario | 0.80 | 0.180 | 0.046 |
| Índice forma foliolo secundario | 0.76 | 0.147 | 0.045 |
σ 2 c Varianza clonal; σ 2 s*c Varianza de sitio por clon
En ensayos clonales de Eucalyptus urophylla S. T. Blake, en el sureste de México, se indican correlaciones genéticas tipo B de moderadas a altas para pares de sitios, de 0.57 a 0.94 para diámetro y de 0.51 a 0.93 para volumen (Sánchez-Vargas et al., 2004). En ensayos de progenie de Acacia mangium Willd. serhalló baja interacción genotipo-ambiente con valores de Bpara volumen de 0.90, diámetro de 0.96 y altura de 0.80 (Pavlotzky y Murillo, 2014).
Parámetros de estabilidad y nivel de interacción de los clones
Para diámetro, altura, volumen e inclinación del fuste, el número de clones que contribuyó significativamente a la interacción genotipo-ambiente fue reducido (Fig. 2), lo que indica que excluyendo pocos clones se reduce significativamente la interacción genotipo-ambiente.
Algunos clones con volumen promedio alto también mostraron alta interacción, lo que se debe a que tienen valores muy altos en un sitio, muy alejados del valor medio, lo que genera alta varianza. Los clones 67, 71, 72, 85, 95 y 120 son los más interactivos en volumen y, en general, para altura y diámetro. El 71 resultó interactivo y además de alta inclinación del fuste, lo que apunta a su eliminación de un grupo de clones superiores para usarse extensivamente. Los clones 67 y 120 son además interactivos para inclinación del fuste, pero de alta rectitud promedio.
Sin embargo, los clones 67, 71, 72 y 95 aunque son interactivos en volumen, no se debieran excluir de un grupo selecto de clones para usarse en el sitio 1 por sus valores altos de volumen en este sitio. El clon 67, por ejemplo, resultó muy interactivo, pero es porque en el sitio 1 presentó un valor superior extraordinario en volumen, en la jerarquización es el número 1, mientas en el sitio 2 es el número 6.
Los clones desarrollados en un país presentarán resultados distintos en otro y, aunque pudiera haber los que tuvieran un comportamiento estable, es necesario evaluarlos en la región de interés.
Impacto de la exclusión de los clones más interactivos
Al excluir del análisis a los clones más interactivos por volumen de poco promedioWi general (120, 85 y 61) y por su valor de ecovalencia ( ), se observó un aumento considerable en la correlación genética tipo B de volumen. La exclusión de clones interactivos no impactó negativamente en la heredabilidad y se obtuvo un ligero aumento en la heredabilidad para las variables de crecimiento en el análisis conjunto (Tabla 5). En el análisis para cada sitio, la heredabilidad se mantuvo en general igual a la estimada con los datos originales o disminuyó ligeramente como en el sitio 2 o aumentó ligeramente en el sitio 1 en las variables de crecimiento.
Tabla 5 Valores de Hi2, Hc2 y rBen análisis conjunto, Hi2 y Hc2 en análisis por ensayo, antes y después de excluir los tres clones más interactivos para la característica de volumen a los siete años en dos ensayos clonales de Cedrela odorata L. ubicados en Santa Genoveva, Campeche (Sitio 1) e Isla, Veracruz (Sitio 2).
| Variable | Análisis con datos originales | Análisis excluyendo tres clones | ||||||||||||
| Análisis conjunto | Sitio 1 | Sitio 2 | Análisis conjunto | Sitio 1 | Sitio 1 | |||||||||
|
|
|
rB |
|
|
|
|
|
|
rB |
|
|
|
|
|
| DIAM | 0.12 | 0.53 | 0.74 | 0.09 | 0.28 | 0.09 | 0.35 | 0.13 | 0.58 | 0.88 | 0.09 | 0.30 | 0.22 | 0.62 |
| ALT | 0.09 | 0.48 | 0.74 | 0.05 | 0.19 | 0.15 | 0.49 | 0.10 | 0.53 | 1.00 | 0.05 | 0.20 | 0.13 | 0.45 |
| VOL | 0.08 | 0.42 | 0.79 | 0.07 | 0.24 | 0.2 | 0.58 | 0.08 | 0.46 | 0.99 | 0.07 | 0.25 | 0.18 | 0.55 |
| INC | 0.09 | 0.48 | 1.00 | 0.10 | 0.31 | 0.06 | 0.27 | 0.09 | 0.48 | 0.94 | 0.10 | 0.31 | 0.07 | 0.29 |
| FFUS | 0.07 | 0.44 | 1.00 | 0.07 | 0.23 | 0.03 | 0.16 | 0.07 | 0.45 | 1.00 | 0.07 | 0.23 | 0.04 | 0.18 |
| TRAM | 0.03 | 0.25 | 1.00 | 0.02 | 0.06 | 0.03 | 0.16 | 0.03 | 0.22 | 1.00 | 0.02 | 0.09 | 0.01 | 0.08 |
| LHOJA | 0.10 | 0.46 | 0.64 | 0.09 | 0.29 | 0.24 | 0.64 | 0.10 | 0.47 | 0.64 | 0.10 | 0.32 | 0.07 | 0.29 |
| AHOJA | 0.08 | 0.39 | 0.53 | 0.07 | 0.23 | 0.24 | 0.64 | 0.06 | 0.32 | 0.53 | 0.07 | 0.25 | 0.24 | 0.63 |
| LPEC | 0.06 | 0.32 | 0.29 | 0.11 | 0.34 | 0.22 | 0.61 | 0.07 | 0.35 | 0.33 | 0.11 | 0.35 | 0.22 | 0.62 |
| LPU | 0.33 | 0.81 | 0.90 | 0.17 | 0.46 | 0.51 | 0.86 | 0.33 | 0.81 | 0.90 | 0.17 | 0.47 | 0.51 | 0.85 |
| NPFS | 0.11 | 0.55 | 0.94 | 0.09 | 0.30 | 0.13 | 0.46 | 0.11 | 0.55 | 0.92 | 0.10 | 0.31 | 0.14 | 0.47 |
| LFS | 0.07 | 0.33 | 0.38 | 0.09 | 0.29 | 0.25 | 0.65 | 0.07 | 0.34 | 0.40 | 0.09 | 0.30 | 0.25 | 0.65 |
| AOFS | 0.25 | 0.72 | 0.80 | 0.15 | 0.41 | 0.41 | 0.80 | 0.26 | 0.75 | 0.84 | 0.15 | 0.43 | 0.41 | 0.80 |
| IFFS | 0.18 | 0.61 | 0.76 | 0.11 | 0.34 | 0.35 | 0.75 | 0.18 | 0.64 | 0.82 | 0.11 | 0.36 | 0.34 | 0.75 |
DIAM = Diámetro (cm), ALT = Altura (m), VOL = Volumen (dm3), INC = Inclinación, FFUS = Forma del fuste, TRAM = Tipo de ramificación, LHOJA y AHOJA = Longitud (cm) y ancho (cm) de hoja, LPEC y LPU = Longitud de peciolo (cm) y de peciolulo (mm), NPFS = Número de pares de foliolos secundarios, LFS, AOFS y IFFS = Longitud (mm), ancho (mm) e índice de forma de foliolo secundario (largo/ancho).
Al excluir tres de los clones interactivos, la correlación B aumentó también en diámetro y altura; respecto a las variables morfológicas también aumentó ligeramente para longitud de peciolo, longitud, ancho e índice de forma de foliolos secundarios (Tabla 5). Las variables que mantuvieron el valor de Bfueron forma del fuste, tipo de ramificación, longitud y ancho de hoja y longitud del peciolulo. Por otro lado, en las variables inclinación y número de pares de foliolos secundarios la correlación disminuyó ligeramente.
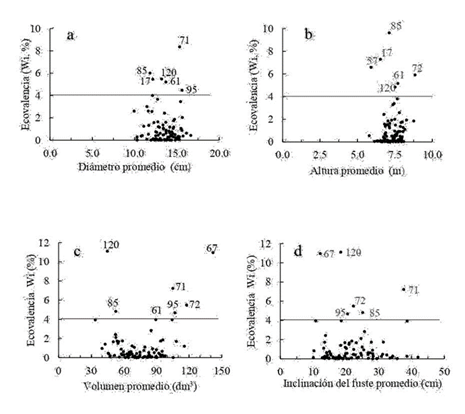
Sobre la línea horizontal en la gráfica se identifican los clones que tienen una contribución a la varianza de la interacción genotipo-ambiente mayor a 4 % en la característica respectiva.
Figura 1 Relación entre la contribución relativa a la varianza de interacción genotipo-ambiente (ecovalencia, Wi en %) y el valor promedio del clon en diámetro normal (a), altura (b), volumen (c) e inclinación del fuste (d) a los siete de edad.
Implicaciones para el mejoramiento genético
Las heredabilidades fueron moderadas; el uso de portainjertos pudo afectar las estimaciones ya que este ejerce influencia sobre el injerto debido a que cada portainjerto es un genotipo diferente. Así el rameto es resultado de la interacción que se da entre ambos, lo que aumenta la variación dentro del clon. La alta mortalidad y heterogeneidad ambiental en el sitio Santa Genoveva generó menor heredabilidad que en Isla. Con menor número de plantas, la variación dentro de clones aumenta, lo que ocurre igualmente con las diferencias ambientales en el sitio de prueba. Para aumentar la estimación del control genético de cualquier característica se debe aumentar el número de rametos por clon en los sitios de evaluación (White et al., 2007).
Las variables tipo de ramificación y forma del fuste son características económicas importantes, pero no presentaron un buen control genético y la estimación de las correlaciones genéticas se dificultó debido a la falta de normalidad de estas variables.
A pesar de la diferencia ambiental entre los dos sitios, se presentó interacción genotipo ambiente moderada. Esta interacción pudo deberse tanto a factores ambientales como a diferencias en el manejo. El ensayo de Santa Genoveva contó con riego en los primeros cinco años, pero con mayor mortalidad, lo que significó menos competencia; así, los clones tuvieron mayor volumen que en el ensayo de Isla. Sin embargo, lo que causa dicha interacción es el número reducido de clones, por lo que excluyendo estos clones superiores se puede obtener menor interacción genotipo-ambiente.
Los clones excluidos del análisis son interactivos para las variables diámetro, altura, inclinación y volumen. Si bien el volumen es una característica económica importante, para el caso de cedro rojo también es importante elegir clones con baja interacción en inclinación. Esta variable es importante, ya que a menor inclinación presentan mejor conformación y es un indicador de que su tolerancia al ataque del barrenador es mayor. Al seleccionar clones sobresalientes en volumen también se mejora la rectitud. Los clones 67, 71, 72 y 95 presentaron alta interacción, sin embargo, presentaron volúmenes promedio altos para el sitio 1 siendo los primeros lugares en la jerarquización. Para no disminuir ganancia genética estos clones pueden ser utilizados solamente en regiones de Campeche, y el resto de los clones puede ser utilizado en una sola zona de mejoramiento considerando ambos lugares de prueba.
Conclusiones
Las variables evaluadas difieren en el grado de control genético, aunque en ambos sitios de prueba las heredabilidades se determinaron bajas, resultaron mayores en el sitio de menor productividad. El número reducido de rametos de cada clon en los ensayos y el uso de patrones para injerto genera alta variación ambiental en relación con la variación genética. Es posible obtener ganancia genética por volumen debido a su coeficiente de variación alto. En ambos sitios, al seleccionar por volumen, se disminuirá la inclinación de los árboles. Algunas variables morfológicas de hojas presentaron correlación genética moderada con las variables de crecimiento. La presencia de interacción genotipo-ambiente es baja y se presenta en pocos clones. La exclusión de algunos clones interactivos disminuye la interacción genotipo-ambiente y aumenta un poco la heredabilidad en el análisis conjunto para variables de crecimiento, sin cambios en los valores estimados de heredabilidad por sitio individual.

