










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkUniversidad y ciencia
versión impresa ISSN 0186-2979
Universidad y ciencia vol.28 no.2 Villahermosa ago. 2012
Artículos
Balance de selenio en corderos suplementados con selenio orgánico
Selenium balance in lambs fed organic selenium
AG López-Gutiérrez1, JE Ramírez-Bribiesca1 *, R López-Arellano2, A Revilla-Vázquez2, J Tórtora-Pérez3, JR Bárcena-Gama1
1 Programa en Ganadería, Instituto de Recursos Genéticos y Productividad, Colegio de Postgraduados, Texcoco, Estado de México. Km 36.5 carretera México-Texcoco. CP 56230, Tel. 01 (595) 95 20 200 Ext.1714. (AGLG)(ERB)(RBG) * Correo electrónico: efrenrb@colpos.mx.
2 División de Farmacéutica, FES-C, Campo I. Universidad Nacional Autónoma de México, México. (RLA)(ARV)
3Facultad de Veterinaria, FES-C, Campo IV. Universidad Nacional Autónoma de México, México. (JTP)
Artículo recibido: 18 de noviembre de 2009,
Aceptado: 09 de enero de 2012
RESUMEN
El objetivo del trabajo fue determinar el balance de Se y la respuesta en la glutatión peroxidasa. Se llevó a cabo un ensayo para evaluar el efecto de la concentración de Se en la dieta, así como la adición de Se orgánico, los tratamientos fueron: T1) Dieta testigo, la cual contenía 0.33 mg de Se y T2) Testigo + 300 mg de Selplex, con una concentración final de 0.93 mg de Se. El ensayo se llevó a cabo usando 8 corderos con un peso inicial de 30.5 ± 2.09 kg, donde la concentración de Se en tejidos, la actividad de glutatión peroxidasa y balance de Se fueron analizados. El Se en la dieta no afecto la concentración de esté en tejidos para el grupo testigo así como para los animales que recibieron Se orgánico. La actividad de glutatión peroxidasa no tuvo diferencias entre tratamientos (p < 0.11) aunque disminuyó ligeramente conforme transcurrieron los días del experimento, tanto en el testigo como en el tratamiento con Se orgánico. Los animales que recibieron Se orgánico tuvieron una mayor (p = 0.054) retención y una mayor (p < 0.05) absorción con relación al grupo testigo. La adición de Se orgánico mejora ligeramente el balance en los animales aunque no se refleja en la actividad de glutatión peroxidasa, ni en la concentración en tejidos.
Palabras clave: Selenio orgánico, tejidos, glutatión peroxidasa, balance de selenio.
ABSTRACT
The purpose of this study was to determine the balance of Se and the glutathione peroxidase activity. A test was carried out to evaluate the effect of the concentration of Se on the diet, as well as the addition of organic Se. Treatments were: T1) control diet, with 0.33 mg of Se and T2) control + 300 mg of Selplex, with a final Se concentration of 0.93 mg. The test used 8 lambs with an initial weight of 30.5 ± 2.09 kg. The concentration of Se in tissues, the glutathione peroxidase activity and the balance of Se were analysed. The Se in the diet did not affect its concentration in the tissues of the control group and in those of the animals that received organic Se. The glutathione peroxidase activity presented no difference among treatments (p < 0.11), although it decreased slightly during the experiment, both in the control group and in the group under treatment with organic Se. Animals given organic Se presented a higher retention ( p = 0.054) and a greater absorption (p < 0.05) compared with the control group. The addition of organic Se slightly improved its balance in the animals, though it was not reflected in the glutathione peroxidase activity nor in its concentration in the tissues.
Key words: Organic selenium, tissues, glutathione peroxidase, selenium balance.
INTRODUCCIÓN
La deficiencia de selenio (Se) es un problema endémico desde el altiplano hasta el sur de México, por lo tanto los alimentos que se producen en esta región son pobres en Se, ocasionando concentraciones inadecuadas para mantener las funciones fisiológicas en los animales. Esta deficiencia puede prevenirse de diferentes maneras, el método de elección depende de las condiciones productivas de los animales (McPherson & Chalmers 1984). Las formas de suministrar Se es a través de mezclas minerales, soluciones inyectables de selenito de sodio, soluciones vía oral, bolos intrarruminales, en el agua ó suplemento mineral a libre acceso (Pérez 2007). Las fuentes de Se permitidas para ganado son en forma de selenito de sodio, selenato de sodio y levaduras con Se (FDA 2005). A partir de 2005, la FDA permite la utilización de levadura con Se en la alimentación de ovinos a una concentración no mayor a 0.3 ppm. La literatura comercial menciona un mejor aprovechamiento de Se cuando es suministrado como selenolevadura, debido a que Sacharomy-ces cerevisiae es un microorganismo con alto valor alimenticio, con contenido adecuado de proteínas, lisina, isoleucina, treonina y por supuesto Se en las formas de selenocistina y selenometionina (Urban 2005).
Específicamente, las bacterias ruminales metabolizan la forma inorgánica, incorporándolo a la proteína microbiana como selenometionina; por lo que un aumento en la retención microbiana puede causar competencia con el animal (Kincard 1995). Después de que el Se pasa los pre-estómagos, ya sea en forma iónica o incorporada a las bacterias, se absorbe en duodeno y se transporta por plasma para incorporarse a los eritrocitos, leucocitos, mioglobina, nucleoproteínas, miosina y varias enzimas (Harrison & Conrad 1984). Las diferencias de absorción entre rumiantes y no rumiantes, se bebe a la interacción del mineral ocasionada por los microorganismos del rumen, que cambian una parte en formas insolubles (Se elemental y selenuros) y otra porción la agregan a proteínas para formar algunos selenoaminoácidos como son la selenometionina y selenocistina (Harrison & Conrad 1984).
La eliminación del Se en rumiantes es en heces y ocurre en dos etapas: La primera está relacionada con la dosis administrada; la segunda es influenciada por la cantidad de Se presente en el animal (Pope et al. 1979). Otra forma de eliminación es en forma de exhalación, excreción urinaria o excreción endógena fecal. En la secreción biliar aproximadamente un 28 % de la ingestión total corresponde a Se; a pesar de que la mayor parte se reabsorbe, el resto se eliminará en las pérdidas endógenas fecales, las cuáles afectan negativamente el balance de Se (Langlands et al. 1986).
Las diferencias en la disponibilidad de Se, depende de la forma en que se suministra; de tal manera que cuando se proporciona en forma de Se inorgánico, una porción se reduce químicamente a formas insolubles por los microorganismos del rumen, lo que disminuye su absorción. El Se inorgánico solo se incorpora dentro de un aminoácido en específico que es la cisteína dentro de la selenoproteína, mientras que la selenometionina puede ser incorporada directamente dentro de la proteína corporal pero sin actividad (Kincard 1995) funcionando como almacén de Se. La forma de suministrar Se orgánico está asociado con levaduras y metionina, se absorbe fácilmente por la pared intestinal (Weiss 2003) y entra al "pool" de metionina, incorporándose a las proteínas, pero ahora como un "pool" para Se orgánico (Suzuki & Ogra 2002). La concentración y la actividad de la glutatión peroxidasa en la sangre, es constantemente sugerida para valorar la concentración de Se de manera indirecta, sin embargo siguen existiendo puntos oscuros en la estabilidad y disponibilidad de la enzima para usarse como diagnostico de Se. El objetivo de este trabajo fue evaluar la respuesta fisiológica de la glutatión peroxidasa y el balance de Se en corderos suplementados con selenio orgánico a través de una levadura.
MATERIALES Y MÉTODOS
Animales y manejo
Se utilizaron 8 corderos machos, criollos Peli-buey, con un peso inicial promedio de 30.5 2.09 kg. Se alojaron enjaulas individuales, equipadas con comedero y bebedero, dentro de la Unidad metabólica ubicada en el Colegio de Postgraduados, Montecillo, Texcoco, Estado de México. Los corderos tuvieron acceso libre al consumo de agua, la ración fue de 1.080 kg de alimento base seca al día, a las 7 y 15 horas, durante 35 d; los primeros 21 d fueron de adaptación y posteriormente los otros 14 d se usaron para obtener muestras de sangre, heces y orina.
Dieta experimental
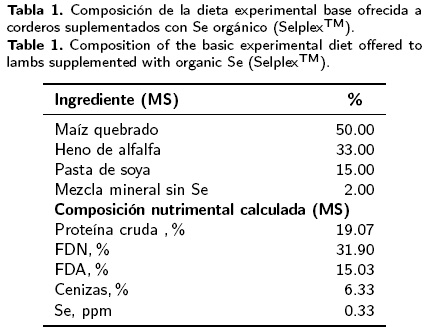
La dieta se calculó de acuerdo a las recomendaciones de NRC (2007), con un contenido de 0.33 ppm de Se, Tabla 1. El Se orgánico (SelplexTM) utilizado contiene una concentración de 2 mg de Se por cada 1000 mg. Los tratamientos fueron: T1) Dieta testigo, la cual contenía 0.33 ppm de Se y T2) Testigo + 300 mg de Selplex, con una concentración final de 0.93 ppm de Se.
Variables de estudio
Actividad de Glutatión Peroxidasa en sangre
Se tomaron muestras de sangre un día antes de que los corderos recibieran los tratamientos y posteriormente se realizaron 4 muestreos en la fase de colección. La sangre se obtuvo en tubos vaccoutainer con anticoagulante, conservándose en hielo y almacenados en el laboratorio de Farmacia de la facultad de Química de la Universidad Nacional Autónoma de México a -20°C hasta su análisis.
Balance de Se
Durante el experimento se utilizó oxido de cromo en la dieta a una concentración de 0.35 % para determinar la producción fecal diaria y el coeficiente de excreción de Se. Las muestras de heces se obtuvieron a las 1:00 y 7:00 hrs post-alimentación y se colectaron diariamente después del día 21. Las muestras de orina se colectaron en los días 21, 28 y 35. Todas las muestras se conservaron a -20°C hasta el análisis de laboratorio. El balance de Se fue calculado con las concentraciones de Se en el alimento, heces y orina; utilizando las siguientes formulas (Koening et al. 1997): Absorción (A): A = I - H y retención (R): R = I - (H + O); donde: I = concentración de Se ingerido, H = concentración de Se en heces y O = concentración de Se en orina.
Obtención de muestras de tejidos
Los animales se sacrificaron en un rastro particular ubicado en San Martín Texmelucan. Antes del sacrificio se pesaron, se desensibilizaron con una pistola de embolo oculto y se desangraron. Posteriormente se tomaron muestras de músculo, hígado, corazón, pulmón y riñón. Estas se conservaron en congelación a -20 C, hasta su análisis.
Análisis de laboratorio
La concentración de cromo en las muestras de alimento y heces se realizó por absorción atómica. En las muestras de alimento, heces, tejidos y orina se determinó el contenido de MS para las dos primeras y la concentración de Se para todas, utilizando un espectrofotómetro de absorción atómica Varian® SpectraAA 800 a través de la metodología descrita por Baeza (2005). La actividad de la enzima glutatión peroxidasa en sangre, se determinó siguiendo la metodología descrita por Sarabia (2004).
Diseño experimental, modelos y análisis estadísticos
Los corderos se distribuyeron de forma aleatoria en dos tratamientos con cuatro repeticiones cada uno. Para la variable concentración de Se en tejidos se utilizó un diseño al azar con 2 tratamientos y 4 repeticiones cada uno. Los resultados fueron analizados con el procedimiento modelo general lineal (SAS 2000). Las diferencias entre medias de tratamientos se realizaron con la prueba de t-Student.
El modelo estadístico utilizado fue:
Yij = μ + Ti + Eij
Donde:
Yij= variable de respuesta correspondiente al i-ésimo tratamiento en la j-ésima repetición; μ = constante que caracteriza a la población; Ti = Efecto del i-ésimo tratamiento, i = 1, 2; Eij= Efecto del error del i-ésimo tratamiento en la j-ésima repetición, j = 1, 2, 3, 4.
Las variables balance de Se y actividad de glutatión peroxidasa, fueron medidas a través del tiempo y fueron analizadas con el procedimiento Mixed de SAS (SAS 2000). El modelo estadístico utilizado fue el siguiente:
Yijk = μ + Ti + Rj + Aij + Pk + (TP)ik + Eijk
Donde:
Yijk = variable de respuesta correspondiente al i-ésimo tratamiento, de la j-ésima repeticion, en el k-ésimo período. μ = constante que caracteriza a la población. Ti = Efecto del i-esimo tratamieto, i= 1, 2; Rj = Efecto de la j-ésima repeticion, j = 1, 2 , 3, 4; Aij= Efecto de la j-ésima repeticion en el i-ésimo tratamiento;Pk = efecto del k-esimo periodo; TPik = efecto de la interaccion del i-ésimo tratamiento por el k-ésimo periodo; Eijk = error experimental.
RESULTADOS
Actividad de glutatión peroxidasa (GSH-Px) en sangre
No se encontraron diferencias en la actividad de GSH-Px entre los corderos del tratamiento testigo y los del tratamiento con Se orgánico (SelplexTM). Sin embargo, en el tercer muestreo del experimento se observó una mayor actividad de esta enzima en los corderos suplementados. Existió una tendencia a disminuir la actividad de GSH-Px conforme transcurrieron los días del experimento, tanto en el testigo como en el tratamiento con SelplexTM (Tabla 2). Con respecto al análisis estadístico en el tiempo se encontró una diferencia altamente significativa (p = 0.0011), también se presentó una interacción entre tratamiento y tiempo (p = 0.08).
Balance de Se
Los corderos que recibieron Se orgánico (SelplexTM) tuvieron una mayor absorción (p < 0.05) y retención (p = 0.054) de Se, en relación a los que se alimentaron con el tratamiento testigo. No se encontró interacción entre el tratamiento y el tiempo para ambos casos (Tabla 3).
Concentración de Se en tejidos
La concentración de Se en los tejidos solo presentó diferencias en el tejido cardiaco (p < 0.1), la desviación estándar fue muy alta. Con respecto a los demás tejidos no hubo diferencias significativas (p > 0.05) entre los tratamientos, Tabla 4.

DISCUSIÓN
Actividad de glutatión peroxidasa (GSH-Px) en sangre
Los resultados encontrados en este experimento coinciden parcialmente con lo encontrado por Van Ryssen et al. (1989) y Gunter et al. (2003) quienes reportan que no encontraron diferencias en la actividad de Glutatión peroxidasa en sangre en ovinos al suplementar Se orgánico (SelplexTM) en la dieta. La actividad de glutatión peroxidasa se utiliza como un indicador del estatus de Se en el animal. Sin embargo, conforme el Se en sangre aumenta, la enzima GSH-PX se satura, por lo que en conjunto, tiende a no incrementar la actividad de la enzima (Juniper et al. 2008a) y no resulta un buen indicador para diferenciar la suplementación de Se. En este punto se concluye que la determinación de glutatión peroxidasa no resulta adecuada para el diagnostico de Se en corderos, debido a su amplia variabilidad y a la falta de asociación con el balance de absorción.
Balance de Se
Se ha reportado un efecto lineal entre la concentración de Se en la dieta y la eliminación, ésta se incrementa conforme aumenta el Se en la dieta. Del mismo modo, hay un efecto lineal con la absorción y con esto, se alcanza un balance positivo (Ivancic & Weiss 2001). Los corderos que recibieron el tratamiento testigo no presentaron un balance positivo de Se, esto significa que el Se fue retenido en el organismo y se liberó menor cantidad que la suministrada (Tabla 3). Las diferencias en el balance de los grupos testigo y suplementado pueden ser parcialmente atribuidas a la influencia de la actividad microbiana o posiblemente a una mayor retención en tejidos y una menor tasa de pasaje de la digesta a lo largo del tracto gastrointestinal (Patterson et al. 1989). La composición de la dieta influye en la disponibilidad de Se para la absorción en rumiantes, además de que es determinante en el entorno químico y las poblaciones microbianas en el rumen (Koening et al. 1997). En dietas altas en concentrado disminuye el pH del rumen y se favorece la conversión de Se a formas no disponibles (Ivancic & Weiss 2001).
En la Tabla 3, se presentan los resultados de los animales que recibieron el tratamiento con Se orgánico (SelplexTM), la concentración de Se tuvo un incrementó en orina, hay muy poca información de la liberación de Se en corderos. Sin embargo, un trabajo asociado es el de Gutiérrez et al. (2005) donde encontró que la concentración de Se en heces fue bajo e infiere que la vía fecal no fue la principal vía de excreción como pudo ser la urinaria, lo cual pudo deberse a que hubo un aporte adecuado de Se. Los corderos que recibieron el tratamiento testigo tuvieron una menor concentración de Se en orina, pero la principal vía de excreción fue en heces. Esto coincide con lo reportado por Krishnamurti (1997) quien menciona que las posibles fuentes de Se en la materia fecal son endógenas e incluyen secreciones biliares, pancreáticas, en saliva, células de la sangre y la movilización en los tejidos. Los precursores de estas secreciones se derivan de la circulación general o de la absorción de Se en el intestino y son nuevamente excretados en el duodeno. La excreción urinaria es un mecanismo primario para disminuir la retención de Se y así, mantener la homeostasis (NRC, 1983).
Concentración de Se en tejidos.
El riñón y el hígado son los tejidos que más acumulan Se, y la acumulación en estos tejidos depende de la deficiencia o el exceso del mineral administrado. El trabajo realizado en esta investigación, la mayor concentración de Se fue en hígado y riñón, específicamente el grupo testigo acumulo mas Se en riñón (p < 0.06), esta información es coincidente con Cousin et al. (1961), quienes reportan mayor concentración de Se en riñón, cuando la concentración en la dieta fue bajo. Los datos reportados por Juniper et al. (2008b) comparando a un tratamiento testigo sin suplemento de Se, contra un tratamiento con Se orgánico utilizando una dosis de 0.3 mg kg-1 de alimento, encontró que la concentración total fue también mayor en el riñón y posteriormente en hígado, corazón y músculo. Esta respuesta es compatible con lo informado por Tylor (2005) cuando ofreció a corderos dietas con 2.9 mg kg-1 de Se orgánico (SelplexTM). Similarmente Quin et al. (2007) reportan una concentración de Se mayor en el riñón, hígado, músculo y tejidos en ovinos con dietas suplementadas con 0.10 mg kg-1 de Se orgánico (SelplexTM). En nuestro estudio se utilizaron concentraciones de 0.3 y 0.9 mg kg-1 resultando adecuadas como suplemento, y además están por debajo del nivel de 5 mg kg-1, considerada como la concentración máxima tolerable (NRC, 2007).
La suplementación de Se orgánico a la dieta de corderos no incrementó la concentración de Se en los diferentes tejidos evaluados, ni modificó la actividad de la enzima glutatión peroxidasa en sangre.
Adicionar Se en forma orgánica (selplex) mejora el balance de Se, aunque no se vio reflejado en la actividad de la glutatión peroxidasa, ni en la concentración de Se en tejidos.
AGRADECIMIENTOS
Esta investigación fue parcialmente financiada por la LPI-7. Inocuidad, calidad de alimentos y bioseguridad. Colegio de Postgraduados.
LITERATURA CITADA
Baeza Hernández MG (2005) Validación de un método es un horno de microondas para la cuantificación de selenio por absorción atómica en materia prima y fluido ruminal. Tesis. UNAM. [ Links ]
Gunter SA, PA Beck, Phillips JM (2003) Effects of supplementary selenium source on the performance of blood measurements in beef cows and their calves. Journal of Animal Science. 81: 856-864. [ Links ]
Gutierrez OC, Spross SA, Rosiles MR, Ducoing WA, Ortiz HA (2005) Selenio sanguíneo y fecal en ovinos a partir de comprimidos inorgánicos intrarruminales. Veterinaria México. 36 (3) 313: 324. [ Links ]
Harrison JH, Conrad HR (1984) Effect of selenium intake on selenium utilization by the nonlactating dairy cow. Journal Dairy Science. 67: 219-223. [ Links ]
Ivancic J, Weiss WP (2001) Effect of dietary sulfur and selenium concentrations on selenium balance of lactating Holstein cows. Journal of Dairy Science. 84: 225-232. [ Links ]
FDA (2005) CVM update. FDA permits the use of selenium yeast in sheep and goat feed. Disponible en: http://fda.gov/cvm/CVM_Updates/SEsheep.htm. [ Links ]
Juniper DT, Phipps RH, Ramos-Morales E, Bertin G (2008a) Effect of dietary supplementation with selenium-enriched yeast or sodium selenite on selenium tissue distribution and meat quality in beef cattle. Journal of Animal Science. 86: 3100-3109. [ Links ]
Juniper DT, Phipps RH, Givens DI, Jones AK, Green C, Bertin G (2008b) Tolerance of rumiant animals to high dose in feed administration of a selenium enriched yeast. Journal of Animal Science. 86: 197-204. [ Links ]
Krishnamurti CR, Ramberg CF, Shariff MA, Boston RC (1997) Compartmental Model Depicting Short-Term Kinetic Changes in Selenium Metabolism in Ewes Fed Hay Containing Normal or Inadequate Levels of Selenium. Journal of Nutrition. 127: 95-102. [ Links ]
Kincard RL (1995) The biological basis for selenium requirements of animals. Journal of Animal Science. 11: 26. [ Links ]
Koening KM, Rode LM, Cohen RDH, Buckley WT (1997) Effects of diet and chemical form of selenium on selenium metabolism in sheep. Journal of Animal Science. 75: 817-827. [ Links ]
Langlands JP, Donald GE, Bowles JE, Smith AJ (1986) Selenium excretion in sheep. Australian Journal of Agricultural Research. 37. 201-209. [ Links ]
MacPherson A, Chalmers JS (1984) Methods of selenium supplementation of rumiants. Veterinary Record. 115.544-546. [ Links ]
NRC (2007) Nutrient Requirements of Small Ruminants. Sheep, goats, cervids, and new world camelids. Editión The National Academy Press, Washington, D.C. 362 p. [ Links ]
Patterson BH, Levander OA, Helzlsouer K, McAdam PA, Lewis SA, Taylor PR, Veillon CI, Zech LA. 1989. Human selenite metabolism: aKinetic model. American Journal of Physiology. 257: 556-567. [ Links ]
Pérez BF (2007) Estudios de perfiles de Se en sangre y líquido ruminal de bolos de selenato de bario. Tesis de licenciatura. FES-C, UNAM. [ Links ]
Pope AL, Moir RJ, Somers M, Underwood EJ, White CL (1979) The effect of sulphur on Se absorption and retention in sheep. The Journal of Nutrition. 109: 1448-1455. [ Links ]
Quin S, Gao J, Huang K (2007) Effects of different selenium sources on tissue selenium concentrations, blood GSH-Px activities and plasma interleukin levels in finishing lambs. Biological Trace Elements. Res. 116: 91-10. [ Links ]
Sarabia Martínez M (2004) Desarrollo de un bolo intrarruminal de liberación prolongada con selenio orgánico de levaduras para bovinos productores de leche. Tesis de maestría. UNAM. [ Links ]
SAS (2000) Statistical Analysis System. The SAS system for Window release 8.0. USA. 558. [ Links ]
Suzuki KT, Ogra Y (2002) Metabolic pathway for selenium in the body: Speciation by HPLC-ICP-MS with enriched Se. Food Addititives and Contaminants 19: 974-983. [ Links ]
Taylor JB (2005) Time-dependant influence of supranutritional organically bound selenium on selenium accumulation in whether lambs. Journal of Animal Science. 83: 1186-1193. [ Links ]
Urban-Sánchez R (2005) Desarrollo de un método para la determinación de selenio componentes presente en una muestra de levadura enriquecida con selenio. Tesis de maestría. UNAM. [ Links ]
Van Ryssen JBJ, Deagen JT, Beilstein MA, Whanger PD (1989) Comparative metabolism of organic and inorganic selenium by sheep. Journal of Agricultural and Food Chemistry. 37: 1358-1363. [ Links ]
Weiss WP (2003) Selenium nutrition of dairy cows: Comparing responses to organic and inorganic selenium forms. En Proc. 19th Alltech Annu. Symp. Nutr. Biotechnology in the Feed and Food Industries. P. Lyons, y K. A. Jacques, Ed. Nottingham University Press, Nottingham, UK. 333-343. [ Links ]

