











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCTION
Tetanus toxin (TT) from Clostridium tetani is a protein that triggers muscular contraction, causing death. TT vaccine is formulated with an inactivated toxin (toxoid) that induces the production of neutralizing antibodies1. Pneumococcal polysaccharides (PPSs) are a complex of carbohydrates present in the capsule of Streptococcus pneumoniae bacteria. The capsule is an important virulence factor that inhibits phagocytosis, promoting infection in the respiratory tract. Pneumococcal vaccine is formulated with a pool of polysaccharides from different serotypes2. S. pneumoniae causes infrequent but severe infections in neonates3.
TT and PPS are two vaccines frequently administered during pregnancy to induce protection in women and newborns4. During intrauterine development, the fetus receives protection by transplacental transfer of immunoglobulin (Ig)G antibodies and, after birth, by maternal lactation containing mainly secretory IgA. Thus, antigen-specific Igs can be measured in serum and secretions such as colostrum and breast milk4.
Colostrum is the primary source of maternal IgA. IgA has two subclasses in humans (IgA1 and IgA2)5. In response to antigenic stimulation, IgA in the mucosa can be produced by T-dependent or T-independent mechanisms6. Ladjeva et al.7 described significant individual variations of IgA1 and IgA2 subclasses in 18 colostrum samples. They analyzed IgA reactivity against polysaccharides and proteins, where IgA2 was more reactive against lipopolysaccharides (LPS); in contrast, the specificity for proteins was mainly found within IgA1.
Several controlled clinical studies have evaluated the effects of antigenic stimulation (vaccine protocols and infectious diseases) during pregnancy on colostrum and milk IgA levels8,9. In a previous study by our group, we found that infectious episodes in the gastrointestinal tract during pregnancy induce higher IgA2 levels in colostrum10. After recurrent gastrointestinal bacterial infections, LPS levels increase in the blood11. LPS is a polysaccharide component of Gram-negative bacteria, and it is known to induce a strong systemic and local immune response by specific antibodies production12.
In this work, we analyzed the effect of antigenic stimulation on the levels of specific IgA subclasses in colostrum. Therefore, the main aim of our study was to evaluate the effects of vaccination or infection during pregnancy on the amount of specific colostrum IgA subclasses in a cohort of Mexican women. The quantitative enzyme-linked immunosorbent assay (ELISA) and Western blot techniques were used to measure colostrum IgA subclasses from 113 women in Mexico City, with an emphasis on IgA1 and IgA2 levels and their reactivity against specific antigens.
METHODS
Calculation of population size (n)
The population size (n) was calculated considering the number of women who received anti-tetanus and anti-pneumococcal vaccines or had infectious episodes of the intestinal tract during pregnancy, 95% confidence interval (Z= 1.96), 5% error.
Population selection
This study included colostrum samples from 113 pregnant, clinically healthy, women (Table 1) who delivered their babies at three large hospitals in Mexico City: Hospital Regional 1º de Octubre ISSSTE (HR 1º October), Hospital Regional de Alta Especialidad de Ixtapaluca 77 (HRAEI), or Hospital de la Mujer (HMuj). All procedures were explained to women, who signed assent and informed consent forms, following the Helsinki Declaration revised in 201313. Clinical information related to the number of infectious episodes of the skin and respiratory, gastrointestinal, or urogenital tracts during pregnancy was obtained from each patient through an interview and clinical examination. Information depicted in the individual clinical files validated the data obtained from the interview at the time of sample collection.
Table 1 Characteristics of the population studied
| Characteristic | Frequencies (%) |
|---|---|
| Age of mothers | Mean = 27.4 ± 4.58 |
| Delivery method | VD = 56 (49.55%) C-section = 57 (50.44%) |
| Number of parturitions | 1st = 108 (95.57%) |
| 2nd = 3 (2.65%) | |
| 3rd = 2 (1.77%) | |
| Hospital | HR 1° October = 35 (30.97%) |
| HRAEI = 40 (35.4%) | |
| HMuj= 38 (33.63%) | |
| Clinical group (Data from documented antigenic stimulation during pregnancy) | Women without documented infectious events = 63 (55.75%) |
| Women with documented infectious events = 50 (44.25%) | |
| Women who received TT vaccination = 27 (23.9%) | |
| Women who received PPS vaccination = 98 (86.72%) | |
| Women with gastrointestinal infection = 13 (11.5%) |
TT, tetanus toxin; PPS, pneumococcal polysaccharide; VD, vaginal delivery; C-section, cesarean section.
Inclusion and exclusion criteria
Colostrum samples were obtained from women between 20 and 35 years of age, with a full-term pregnancy (38-42 weeks), with vaginal delivery (VD) or cesarean section (C-section), and, mainly, singleton births. Exclusion criteria were the presence of chronic maternal disease, previous abortions, preterm pregnancy, multiparous parturition, women with hormonal treatment, or acute disease within 3 weeks before delivery.
Sample processing
Colostrum samples were collected from clinically healthy women between 1 and 12 h post-delivery and frozen at −20°C until use. After defrosting at room temperature, all samples were centrifuged at 2000 g for 10 min at 4°C (Allegra™ X-22R Benchtop Centrifuges Beckman Coulter Life Sciences, Indianapolis, IN, USA) to prepare the corresponding dilutions (see below).
ELISA for quantification of specific Igs
A modification of the quantitative ELISA previously reported10 was used to quantify IgA1 and IgA2 concentrations against TT, PPS, or LPS in colostrum. Anti-TT antibodies quantification: ninety-six-well plates (Immuno Plate MaxiSorp, Thermo Scientific, NY, USA, Cat. Num. 1132249) were coated with 50 µL of 0.5 µg/mL of antigen (TT from C. tetani, Sigma-Aldrich®, Darmstadt, Germany, Cat. Num. T3194-25UG) in borate buffer. Plates were incubated overnight at 4°C. Plates were washed with 100 µL of 0.05% polyoxyethylene sorbitan monolaurate (Tween 20, Sigma-Aldrich Company®, USA, Cat. Num. 9005-64-5) in phosphate-buffered saline (PBS) (PBS-T) for 8 times. The non-specific binding free sites were blocked with 100 µL of 5% skim milk formula (Svelty® diluted in PBS) by incubation during 60 min at 37°C. Anti-PPS antibodies quantification: plates were coated with 50 µL of 5 µg/mL of total antigen (capsular polysaccharides from 23 pneumococcal serotypes, Pulmovax®, Schering-Plough®, West Point, PA, USA, Lot. Num. R033367) in PBS; plates were incubated during 5 h at 37°C and then overnight at 4°C. Plates were washed with 100 µL of PBS-T for 5 times. The non-specific binding free sites were blocked with 5% of bovine serum albumin (BSA, Sigma-Aldrich Company®, USA, Cat. Num. 9048-46-8) with serum fetal bovine (SFB, Biowest®, Riverside, MO, USA, Cat. Num. US1520) diluted in PBS-T (200 µL per well) by incubation during 60 min at 37°C. Anti-LPS antibodies quantification: plates were coated with 50 µL of 5 µg/mL of antigen (LPS from Escherichia coli, Sigma-Aldrich®, Darmstadt, Germany, Cat. Num. L2143) in PBS. Plates were incubated overnight at 4°C and washed with 100 µL of PBS-T for 5 times. The non-specific binding free sites were blocked with 5% BSA diluted in PBS-T (200 µL per well) by incubation for 30 min at 37°C.
From this point, all plates were processed under the same conditions: centrifuged samples were processed by 10-fold serial dilutions starting at ratios of 1:10, 1:100, 1:1000, and 1:10,000. For standard curves, 50 µL of standard serum dilutions from healthy individuals vaccinated against TT, PPC, or reference human serum to LPS were added and incubated for 2 h at 37°C. For the detection of specific IgA subclasses, 50 µL per well of biotinylated antibodies anti-IgA1 (monoclonal mouse antibody, Abcam®, Cat. Num. ab99796; 1:2500) or anti-IgA2 (monoclonal mouse antibody, Abcam®, Cat. Num. ab128731; 1:2000) were added and incubated under the same experimental conditions. Then, 50 µL of streptavidin-horseradish peroxidase (HRP) complex (Streptavidin HRP, Abcam®, Cat. Num. ab7403) dilution were added and incubated for 1 h at 37°C. The presence of labeled antibodies was revealed using 50 µL per well of chromogenic substrate 3’3’’-5-5-tetramethylbenzidine (TMB ELISA Substrate, Abcam®, Cat. Num. 171523) and the reaction was stopped by adding 50 µL of 0.2 M sulfuric acid per well. The absorbance was measured in a Microplate ELISA plate reader spectrophotometer (Sunrise, Tecan® TX, USA) at 450 nm. IgA1 and IgA2 standard curves were obtained for each plate to determine each sample concentration by absorbance interpolation and dilution factor. Results were finally expressed in nanograms of each IgA subclass per milliliter of colostrum (ng/mL).
Western blot
The ability of colostrum to recognize protein antigens from E. coli was evaluated by Western blot. For these assays, 10 representative colostrum samples were evaluated. E. coli O157:H7 from the microbial collection of the National School of Biological Sciences-IPN (Mexico) was grown overnight in LB medium without antibiotics. Bacteria were lysed for 30 min at 4°C with RIPA buffer (20 mM Tris-HCl pH 7.5, 150 mM NaCl, 1 mM Na2 EDTA, 1 mM EGTA, 1% NP-40, 1% sodium deoxycholate, 2.5 mM sodium pyrophosphate, 1 mM b-glycerophosphate, 1 mM Na3VO4, and 1 µg/ml leupeptin). The lysate was then centrifuged at 12,000 ×g for 15 min at 4°C. The supernatant was separated and quantified by the Lowry method, 6X Laemmli buffer was added to the proteins, and samples were heated at 95°C for 5 min. Two-hundred micrograms of proteins were loaded and separated in 10% polyacrylamide preparative gel. The proteins were transferred to a nitrocellulose membrane (0.45 µm; BIO-RAD Cat. Num. 1620115) by a humid chamber (BIO-RAD Mini Trans-Blot® Cell Cat. Num. 1703930) at 100 volts for 1.5 h14. Non-specific binding free sites in the membrane were blocked with 5% skim milk formula (Svelty® diluted in PBS) by incubation during 60 min at 37°C. Colostrum dilutions (1:1000) in 0.05% PBS-T were used as the primary antibody, and the membrane was incubated overnight at 4°C. Biotinylated antibodies such as anti-total IgA (rabbit polyclonal antibody, Abcam®, Cat. Num. ab97218; 1:5000), anti-IgA1 (Abcam®, Cat. Num. ab99796; 1:2500), or anti-IgA2 (Abcam®, Cat. Num. ab128731; 1:2000) in 0.05% PBS-T were added and incubated 1 h at room temperature. Streptavidin-HRP was used at 1:8000 dilution and incubated 1 h at room temperature. Clarity Western ECL (luminol and peroxide) was used as a substrate. Membranes were documented using ChemiDoc™ MP Imaging System with Image Lab™ Software.
Statistical analysis
The IgA concentration distribution was analyzed by the Kolmogorov–Smirnov test. A comparison of Ig concentrations between samples was performed by non-parametric Mann–Whitney U-test or Kruskal-Wallis test. All statistical tests and graphics were developed using statistical program GraphPad Prism (GraphPad® Software version 5, La Jolla, CA, USA).
RESULTS
Population data
The mean age of mothers was 27.4 ± 5.2 years; 50% had a cesarean delivery (C-section) and the other half a VD. For 95% of women, this was their first child. Information on previous infectious episodes and vaccine protocols during pregnancy is shown in Table 1.
Levels of IgA subclasses in colostrum against studied antigens
The levels of IgA subclasses against TT, PPS, and LPS were quantified in colostrum. None of the colostrum Ig levels had a normal distribution. As in the previous reports7, we found that colostrum had higher amounts of IgA1 against TT and, in contrast, IgA2 was more abundant against PPS and LPS (Fig. 1).

Figure 1 Immunoglobulin (Ig)A subclasses in the function of the type of antigen. The bar graph shows the concentrations of IgA1 (blue) and IgA2 (red) against tetanus toxin, Escherichia coli lipopolysaccharide, and Pulmovax 23® (pneumococcal polysaccharides) (n = 113). All values are represented as mean ± standard deviation. All data are expressed in nanograms of Ig/milliliter of colostrum (ng/mL). Statistical analysis was performed using the Mann–Whitney U-test. ***p < 0.001.
IgA subclasses against E. coli antigens by Western blot
The ability of colostrum to recognize protein antigens from E. coli was evaluated by Western blot. For these assays, 10 representative colostrum samples were evaluated. The pattern of proteins recognized by total IgA was similar among the different colostrum samples, showing variations in the intensity of the bands between the different samples (Fig. 2). When IgA1 recognition was evaluated, we found that the banding pattern changed considerably; although some samples varied in the number of proteins recognized, and in the intensity of the bands, the pattern was considerably different when compared with total IgA. In sharp contrast, IgA2 recognized fewer antigen proteins with reduced intensity. The pattern and the intensity of the bands recognized by IgA2 differed from that seen with IgA1 and total IgA.

Figure 2 The SDS-PAGE analysis of colostrum immunoglobulin (Ig)A subclasses against Escherichia coli antigens. Colostrum samples were analyzed to determine the recognition profile of E. coli O157:H7 proteins. The membranes were developed with anti-total IgA (A), anti-IgA1 (B), and anti-IgA2 (C). MW markers, for size comparison, are shown at the left. Lanes 1-10 contain 10 selected colostrum samples.
IgA subclasses did not show variations among population descriptors
Many descriptors played a pivotal role in identifying the characteristics of the population analyzed. The following descriptors were included in the function of the size of the sampled population: the hospital where the mothers were recruited (Supplementary Fig. S1a) and the delivery method (Supplementary Fig. S1b). These comparisons did not show any statistical differences between groups. Hence, we concluded that Ig levels were independent of the descriptors used to stratify this population.
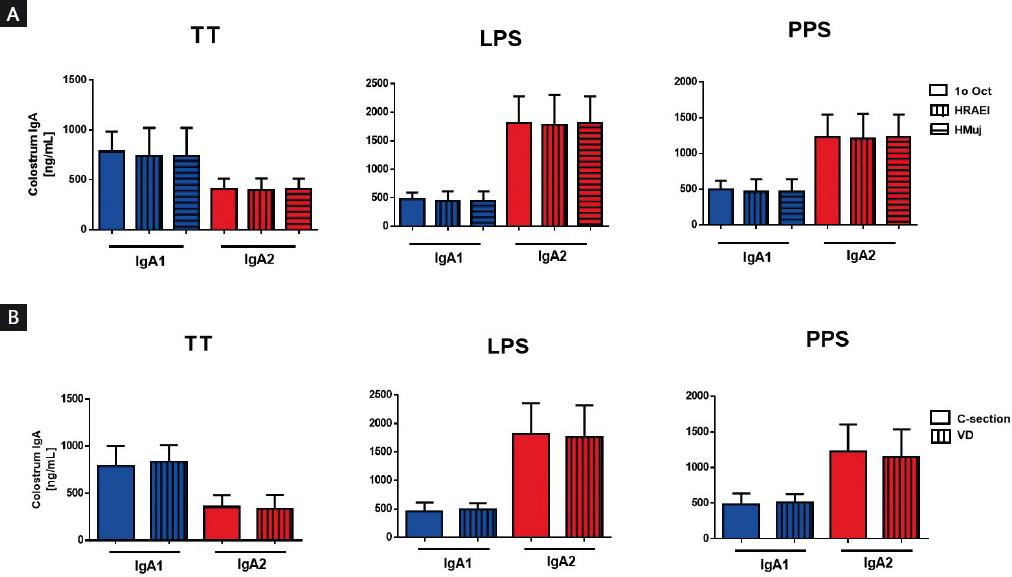
Figure S1 Stratification of colostrum immunoglobulin (Ig) levels in the function of population descriptors. The bar chart shows comparative amounts of IgA against specific antigens in colostrum in the function of (A) hospitals: HR 1° October, HRAEI, and HMuj; and (B) delivery methods: vaginal delivery and cesarean section. Results are shown as mean ± standard deviation. Statistical analysis was performed using the Kruskal-Wallis test for non-parametric independent data.
The concentrations of colostrum IgA subclasses vary in women with previous antigenic stimulation during pregnancy
The amount of colostrum IgA subclasses was compared between women without vaccination or previous infection reports during pregnancy. Results showed that both IgA subclasses against TT, PPS, or LPS increased in women vaccinated with specific antigens (TT or PPS) or women that had infections at the gastrointestinal tract during pregnancy, in comparison with women without infections (Fig. 3). These results were compared with total IgA subclasses levels in the colostrum (Supplementary Fig. S2). This effect was analyzed by the Pearson’s correlation test to evaluate whether multiple interactions with the same antigen during pregnancy had an effect on IgA subclasses levels in colostrum. Although we did not find a statistical correlation, levels of IgA1 tended to increase in colostrum from women with more than one TT vaccination dose. In contrast, IgA2 increased in colostrum from women with multiple episodes of gastrointestinal tract infections during pregnancy when LPS was used as an antigen in the test (Supplementary Fig. S3). As it was already mentioned, 98 women vaccinated with Pulmovax® developed a strong response against polysaccharide, with predominant recognition by IgA2 (Fig. 3).
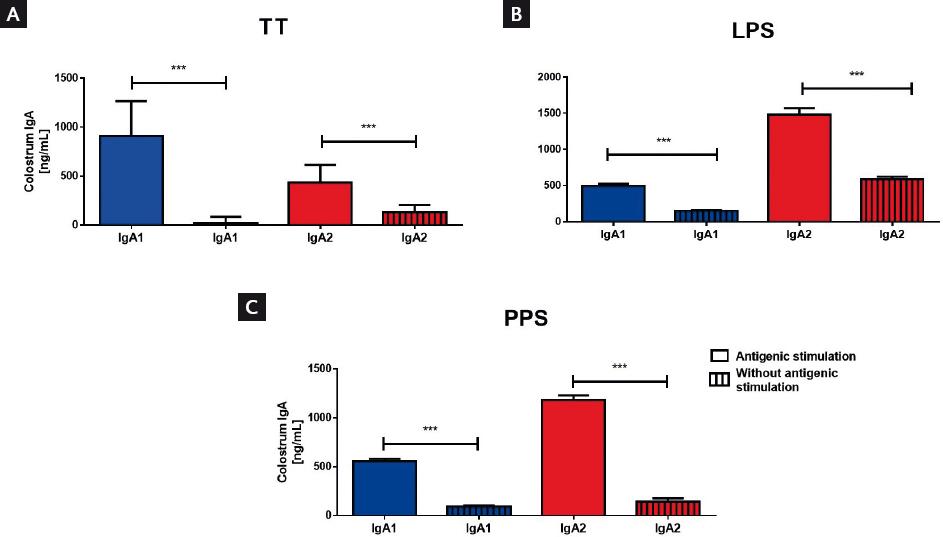
Figure 3 Distribution of immunoglobulin (Ig)A subclasses in the function of the type of antigen, comparing women with or without previous specific antigen stimulation. The amounts of Ig from women with previous infections at the gastrointestinal tract (lipopolysaccharide [LPS]) or vaccination during pregnancy (tetanus toxin [TT] or pneumococcal polysaccharides [PPS]) are compared with each IgA subclass. The bar graphs show IgA1 (blue) and IgA2 (red) between women without documented stimulation during pregnancy (bars with rods n = 63) and women with at least one antigenic stimulation during their pregnancy (bars without rods = 50): TT (n = 27) or PPS (n = 98) vaccination or LPS (n = 13). All values are represented as mean ± standard deviation. All data are expressed in nanograms of Ig per milliliter of colostrum (ng/mL). Statistical analysis was performed using the Mann–Whitney U-test. ***p<0.001.

Figure S2 Quantification of the total immunoglobulin (Ig)A subclasses present in colostrum. Comparative amounts of IgA types in colostrum (IgA, IgA1, and IgA2), n = 113. Bars indicate mean ± standard deviation. All data are expressed in milligrams of Ig per milliliter of colostrum (mg/mL).

Figure S3 Comparison of immunoglobulin (Ig)A1 and IgA2 levels in colostrum against (A) tetanus toxin (TT) or (B) lipopolysaccharide (LPS), in function of the number of vaccination shots (TT) or infections at the gastrointestinal tract (LPS) during pregnancy. All values are represented as mean ± standard deviation. All data are expressed in nanograms of Ig per milliliter of colostrum (ng/mL).
DISCUSSION
According to this study, we found that the previous antigenic exposures during pregnancy affected specific IgA1 and IgA2 concentrations7. However, the mechanisms involved to explain these differences were not studied. As seen from our results, the production of IgA subclasses depends on the chemical nature of the antigen. The humoral responses against most proteins require the interaction of B cells with helper T cells in a thymus-dependent (TD) fashion. B-cell activation by TD antigens requires, among many factors, the interaction between CD40 on B-cells and CD40L on activated Th cells in the germinal centers of the lymph nodes15.
On the other hand, a typical abundant polysaccharide (as in the capsule of Gram-positive bacteria or the external membrane of Gram-negative bacteria) is made up of repeating sequences of few simple monosaccharides, which means that there are multiple copies of identical antigenic determinants. When specific naïve B-cells come in contact with such antigens, these antigenic determinants bind the surface IgM and IgD receptors. This binding leads to the clustering of surface Igs, which generates a signal strong enough to activate the naïve B-cells. Another way to activate B-cells is by the clustering of repetitive epitopes and the stimulation by TLR signals, although in the case of human B-cells, they have low levels of TLR4 on their surface16. T-independent activated B-cells produce and release IgM and “natural” IgA17. The previous reports have suggested that the cells producing IgA2 are mainly found in the gastrointestinal tract, and these cells are predominantly activated through T-independent mechanisms18,19.
A remarkable result, derived from the recognition of E. coli proteins by IgA subclasses, showed (as expected) variations between different colostrum samples, but interestingly, also exhibited differences between IgA1, IgA2, and total IgA recognition. To get a more interesting contrast, when IgA1 mainly recognized the proteins from the lysates, IgA2 displayed a more pronounced reactivity against LPS, contrasting the nature of TD versus TI antigen stimulation in the production of IgA subclasses.
During the final trimester of pregnancy, these IgA1 or IgA2 producing cells migrate to mammary glands under hormonal regulation20 and the expression of different adhesion molecules21,22. These cells can then reside in the mammary gland until lactation23,24, where they produce high levels of IgA subclasses into the mammary acini.
This study has some limitations. It is based on previous antigen stimulation that women recalled and reported during pregnancy. For the correlations, vaccinations (reported in the clinical files) are more reliable parameters for the analysis. Thus, a more systematic clinical study is necessary to demonstrate a correlation between clinical variables and their role in the maternal transference of specific IgA subclasses in mothers and babies. Despite the limitations described above, the results derived from this research provide data and additional information that contribute to solve the controversial findings about the effect of infections during pregnancy on IgA production in the colostrum. This study provides pieces of evidence to understand the effect of infections on the production of IgA subclasses during pregnancy. Moreover, the results offer experimental confirmation in which interactions with specific antigens during pregnancy modify the levels of specific IgA1 and IgA2 in colostrum.

