











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El género Crotalaria incluye entre 600 y 700 especies, 79 de ellas son nativas de América; 21 de México, y nueve de ellas son endémicas (Soto-Estada, 2004). Entre las especies con uso antrópico están C. incana L., C. juncea L., C. longirostrata Hook. & Arn. y C. pumila Ort. como fuente de materia orgánica y N en la agricultura (Jiménez et al., 2005; Basurto-Peña, 2011; Linares & Bye, 2015; Camarillo-Castillo & Mangan, 2020). Crotalaria juncea se utiliza para elaborar cuerdas, cordeles, redes para pesca y papel (Tripathi et al., 2012) y C. retusa L., C. prostrata Roxb. y C. medicaginea Lam. se utilizan en herbolaria por sus efectos antimicrobianos, antiparasitarios y antioxidantes (Devendra et al., 2012).
Crotalaria longirostrata (chipilín) se distribuye en Centroamérica y México, en los estados de Chiapas, Chihuahua, Colima, Guerrero, Jalisco, Estado de México, Michoacán, Morelos, Nayarit, Oaxaca, Puebla, Sinaloa, Tabasco y Veracruz (Soto-Estada, 2004). El hábitat en el que crece la especie es bosque de pino-encino, en áreas abiertas, áreas con maleza y en zonas con cultivos diversos, como frijol (Phaseolus vulgaris L.) y maíz (Zea mays L.), en altitudes entre nivel del mar y 2 300 m (Tapia, 2000; McClung de Tapia et al., 2014). En esas regiones, desde época prehispánica, los brotes tiernos, las hojas y los tallos jóvenes de chipilín se utilizan en la preparación de alimentos y bebidas; asimismo, se utilizan en herbolaria y como forraje (Bye, 1981; Morton, 1994; Centurión et al., 2003; Chávez et al., 2009; Basurto-Peña, 2011; Laguna, 2016). Las hojas son fuente de carotenoides, vitamina C, hierro, calcio y proteínas; además, se ha documentado la presencia de compuestos fenólicos, como flavonoides, saponinas, cumarinas, taninos, antraquinonas, antronas y alcaloides (Arias et al., 2003; Bibi et al., 2015; Cruz-Rodríguez et al., 2017). Además, existen estudios que demuestran mayor diversidad de usos; un caso es el de los extractos de sus tallos, ramas y raíces que inhiben el desarrollo del micelio y la esporulación de fitopatógenos y fitoparásitos como Fusarium sp., Aspergillus flavus Link (Cruz-Rodríguez et al., 2017; Miranda-Granados et al., 2018; Cruz-Rodríguez et al., 2020) y nematodos (Villar & Zavaleta-Mejía, 1990). Todas las estructuras de la planta se han evaluado en nuevas aplicación en herbolaria (Fischler & Wortmann, 1999; Arias et al., 2003; Basurto-Peña, 2011), como abono verde (Desaeger & Rao, 2000), forraje (Arias et al., 2003), alimento para el cultivo y multiplicación de tilapia nilótica (Oreachromis niloticus L.) (Guerra-Centeno et al., 2016) y caracoles de agua dulce (Pomacea flagellata Say) que son alimento, rico en proteína para humanos (Martínez, 2016). Los usos tradicionales de chipilín, los actuales y los documentados como potenciales indican que su demanda tendrá soporte con su cultivo comercial. Lo que también asegura la disponibilidad de plantas para los consumidores regionales y externos y en diferentes épocas del año (Camarillo-Castillo & Mangan, 2020). Un obstáculo para el cultivo de las especies de Crotalaria es la falta de semilla comercial (Vinicius de Araujo et al., 2018). Aunque las semillas para el cultivo pueden obtenerse de poblaciones silvestres, la producción comercial se asegurará con el cultivo de las plantas. Como en el caso de C. brevidens y C. ochroleuca (Mwakha et al., 2020), para conocer el potencial agronómico y la viabilidad comercial de C. longirostrata es necesario conocer y mejorar el rendimiento de las estructuras de interés y promover la producción de semillas de calidad. Esto puede lograrse mediante la domesticación, el cultivo, selección intensiva y el mejoramiento. Sin embargo, los programas de mejoramiento son exitosos cuando cuentan con variabilidad genética del germoplasma, ya que es la base para la selección (Peña-Valdivia et al., 2012; Mwakha et al., 2020). En el caso de C. longirostrata la información disponible sobre sus atributos genéticos y agronómicos es mínima.
La producción y el rendimiento de semilla son resultado de las interacciones entre el genotipo (Mwakha et al., 2020) y el ambiente, que incluye factores como la radiación solar, la temperatura, la disponibilidad de humedad y de nutrientes en el suelo (Rouached & Rhee, 2017). Al respecto, las plantas asimilan macronutrientes, como N, K, P, Ca, Mg, S, y micronutrientes, como Fe, Mn, Cu y Zn, del suelo a través de sus raíces, con lo que garantizar las funciones celulares (White & Brown, 2010). Las alteraciones en la disponibilidad de nutrientes minerales afectan el crecimiento; es el caso de la deficiencia de P, que disminuye el crecimiento de la raíz de Arabidopsis; en contraste, la de Fe o S lo promueve, aunque los tres, junto con el N, son indispensables para la fotosíntesis y la respiración (Rouached & Rhee, 2017). La deficiencia combinada de P y N disminuye la productividad de las plantas silvestres en su ambiente natural y los cultivos (White & Brown, 2010). Esto es porque, el P estimula el desarrollo de los tejidos meristemáticos, el crecimiento de la planta, la conductibilidad hidráulica de las raíces, y sus efectos significativos, junto con el del N, en la altura de la planta, el número de hojas, el número de ramas, el número de frutos y el peso de 1000 semillas de C. brevidens y C. ochroleuca permitió demostrar que el nivel óptimo difiere entre las especies (Mwakha et al., 2020). La producción de semilla se relaciona con el crecimiento de la planta completa; por lo que, para describir el rendimiento de las semillas se necesita evaluar el resto de la planta (Hall, 1980). Los componentes del rendimiento en las plantas involucran caracteres cuantitativos, como número y biomasa de sus órganos individuales, e.g., número o biomasa de ramas, hojas, racimos, frutos y semillas, entre otros (Patrick & Colyvas, 2014). Por ejemplo, el desarrollo de las ramas laterales aumenta el sitio para el desarrollo de frutos; esto se ha demostrado mediante eliminación de la dominancia apical e inducción del desarrollo del número de ramas y el rendimiento de diversos cultivos (Tripathi et al., 2013).
La comparación de los componentes del rendimiento permite el uso y rescate de la diversidad de especies silvestres con valor antrópico, como C. longirostrata, que se han incorporado y conservado en la agricultura tradicional y que están estrechamente relacionados con factores socio-culturales (Peña-Valdivia et al., 2012; Solís Bonilla et al., 2017; Castiñeiras et al., 2000; Rojas-García et al., 2021). La información de la fenología, morfología y rendimiento de las especies silvestres permite contrastar el desarrollo de las plantas in situ y ex situ, diseñar programas de conservación, propagación y cultivo de especies silvestres o semi-domesticadas (Peña-Valdivia et al., 2012). Sin embargo, no se cuenta con esa información de las poblaciones silvestres de C. longirostrata. El rendimiento de semilla y de ciertas estructuras de la planta se ha documentado con fines agrícolas en especies como C. juncea, C. brevidens y C. ochroleuca (Jacobi et al., 2005; Tripathi et al., 2013; Meagher et al., 2019; Rojas-García et al., 2021). Tripathi et al. (2013) evaluaron el efecto del espaciamiento, de 30 x 10 cm a 45 x 20 cm, entre las plantas, y la eliminación de la dominancia apical (por corte del ápice) en los componentes del rendimiento de C. juncea; los autores observaron el incremento de ramas primarias y secundarias por planta (de 7 y de 13 a 17), del número de frutos por planta (en promedio de 47 a 74) y su biomasa (15.5 a 25 g), del número de semillas por fruto (7 a 9) y su rendimiento por planta (9.6 a 15 g) con la separación de las plantas. Los mismos autores observaron que, con excepción del número de semillas por fruto (7.5 a 8), esos componentes del rendimiento incrementaron mediante corte del ápice del tallo. La calidad agronómica de 13 recolectas de C. brevidens y 16 de C. ochroleuca se evaluó en Kenia mediante el cultivo en campo; la variabilidad en anchura de los frutos (1.85 a 1.04 cm y 2.20 a 1.25 cm) y el peso de 1000 semillas (5.44 a 7.70 g y 5.68 a 7.84 g) fue significativa dentro y entre especies. En contraste las diferencias en la longitud de los frutos (en promedio 5.3 cm), el número de estos por planta (14 en promedio) y el número de semillas (83 en promedio) no fueron significativas dentro o entre especies (Mwakha et al., 2020).
El objetivo de este estudio fue determinar el rendimiento de semilla y otros componentes del rendimiento, como fueron número de: ramas, racimos, frutos y semillas por planta, frutos por racimo y por rama, semillas por racimo, por rama y por fruto de plantas silvestres de C. longirostrata y su relación con el sitio de recolecta. La hipótesis es que, los componentes del rendimiento de plantas silvestres de C. longirostrata dependen del ambiente del sitio de recolecta.
Materiales y métodos
Localidades de recolecta e identificación del material vegetal
Plantas de C. longirostrata (Fig. 1) con frutos maduros (color castaño-amarillento, que al agitarse emitían sonido similar de un sonajero cascabel) se cosecharon aleatoriamente, en enero de 2019, de rodales silvestres en las localidades Las Vigas, Ometepec y Llano Grande de los Hilarios Igualapa en Guerrero, México (Tabla 1). Los rodales tenían en promedio 43 ± 3.27 plantas y ocupaban un área aproximada de 100 m2; las plantas tenían altura promedio de 176 ± 0.052 cm y diámetro en la base del tallo de 9.060 ± 0.44 mm. Las plantas fueron identificadas in situ por comerciantes-recolectores de C. longirostrara de cada localidad. La identidad taxonómica se confirmó en ejemplares recolectados ex profeso, herborizados y registrados en el Herbario-Hortorio CHAPA, del Colegio de Postgraduados (accesión Núm. 155775). Los ejemplares de C. longirostrata Hook. & Arn. consultados corresponden a las colectas Núm.: Es-604 de Campeche, 12, 26, 33, 40, 64, 81, 3661, 2768 y 5428 de Chiapas, 14, 3728, 4001 y 4464 de Jalisco, 69, 268 y 2294 de Michoacán, 121 y 530 de Oaxaca, 2036 de Sinaloa y 1636 de Tabasco.
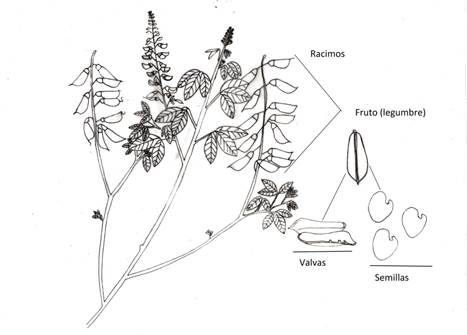
Fig. 1 Estructuras de las plantas de Crotalaria longirostrata Hook. & Arn. recolectadas en Las Vigas, Ometepec y Llano Grande de los Hilarios Igualapa en Guerrero, México.
Tabla 1 Localidades y características de los sitios de muestreo de Crotalaria longirostrata Hook. & Arn. En Guerrero, México (García, 2004; CONAGUA, 2021).
| Localidad | Las Vigas | Ometepec | Llano Grande de los Hilarios Igualapa |
|---|---|---|---|
| Descripción (denominación) | Con manejo agrícola y ganadero (MAG) | Con actividad urbana (AU) | Sin actividades antrópicas (SAA) |
| Ubicación | 16º 40’ 264” N, 98º 27’ 502” O | 16º 41’ 269” N, 98º 24’ 733” O | 16º 45’ 089” N, 98º 25’ 918” O |
| Altitud | 264 m s.n.m. | 350 m s.n.m. | 778 m s.n.m. |
| Clima | Aw2(w) Cálido subhúmedo, lluvias y canícula en verano | Aw2(w) Cálido subhúmedo, lluvias y canícula en verano | A©m(w) Semicálido-húmedo, lluvias en verano |
| Precipitación media annual | 1100 mm | 1100 mm | 1000-1500 mm |
| Temperatura media annual | 25 - 28 °C | 26 - 28 °C | 18 - 26 °C |
| Vegetación aledaña | Árboles de Gliricidia sepium (Jacq.) Kunth ex Walp. | Árboles de Gliricidia sepium (Jacq.) Kunth ex Walp., Guazuma ulmifolia Lam., y herbáceas Porophyllum ruderale (Jacq.) Cass y Euphorbia hirta L.) | Árboles de Ebenopsis ebano, Gliricidia sepium (Jacq.) Kunth ex Walp., Guazuma ulmifolia Lam. Entre otros. |
Variables evaluadas
Plantas
Las plantas se trasladaron, separadas unas de otras, al laboratorio de Biofísica y Fisiología Ambiental Vegetal, del Programa de Posgrado en Botánica del Colegio de Postgraduados. Con base en lo fundamentado por Tripathi et al. (2013), Huerta-Lovera et al. (2018), Mwakha et al. (2020) y Romero-Sánchez et al. (2020) en el laboratorio se evaluó el número de: ramas, racimos, frutos y semillas por planta, frutos por racimo y por rama, semillas por racimo, por rama y por fruto, y el rendimiento de semilla por planta. El diámetro, la longitud y biomasa de los frutos y semillas de una localidad también se determinaron para complementar la información de los componentes del rendimiento. El diámetro y la longitud de los frutos se determinó con un vernier digital (Truper, CALDI-6MP; 14388). La biomasa (g) de los frutos, las valvas y las semillas se determinó en una balanza analítica (Scientech® SA 120, precisión de ± 0.0001 g). Los frutos se mantuvieron en bolsas de papel, dentro de cajas de cartón durante su evaluación.
Suelo
De acuerdo con la NOM.021-RECNAT-2002 (SEMARNAT, 2002), de cada sitio se obtuvieron tres muestras de 1 kg de suelo, de 0 a 20 cm y 20 a 40 cm de profundidad. El suelo se clasificó con los criterios de la WRB (IUSS Working Group WRB, 2015). El análisis de las muestras de suelo, de acuerdo con lo descrito por van Reeuwijk (2002), incluyó: color (carta de colores Munsell), densidad (probeta), retención de humedad (olla de presión), conductividad eléctrica, capacidad de intercambio catiónico, pH, bases intercambiables (Ca++, Mg++, K+) (acetato de amonio), materia orgánica (Walkley-Black), nitrógeno total (micro-Kjeldalhl) y P soluble (Olsen y Dean, 1965).
Diseño experimental y análisis estadístico
El estudio se desarrolló mediante un diseño experimental completamente al azar con tres tratamientos o localidades de muestreo, una planta como unidad experimental y tres repeticiones por localidad. Las variables se analizaron mediante ANDEVA y prueba de comparación múltiple de medias de Tukey (α = 0.05). Cuando no se cumplieron los supuestos normalidad, homogeneidad de varianzas e independencia se empleó la prueba de Kruskal-Wallis (H) seguida de la prueba comparaciones de pares con el programa estadístico InfoStat versión 2016 (Di Rienzo et al., 2008). Los datos que no cumplieron los supuestos de la prueba de Kruskal-Wallis (H) (homogeneidad de varianzas e independencia) se analizaron con la prueba U de Mann-Whitney (P ≤ 0.05). También se aplicó el análisis de correlación de Pearson y de componentes principales (CP) a los componentes del rendimiento con Rstudio 3.6.3. (RStudio Inc, Boston, EE.UU.; RStudio Team, 2020).
Resultados
Componentes del rendimiento
Número de ramas
El número promedio de ramas por planta varió (P ≤ 0.05) entre las localidades. En la localidad MAG (con manejo agrícola y ganadero aledaño) el número de ramas por planta representó hasta 45 % más que en las otras localidades (Tabla 2).
Tabla 2 Componentes del rendimiento (promedio ± E.E.) de plantas de Crotalaria longirostrata Hook & Arn. recolectadas en tres localidades de Guerrero, México.
| Localidad* | Ramas | Racimos | Frutos | Frutos | |
|---|---|---|---|---|---|
| Núm./planta | Núm./racimo | Núm./rama | |||
| MAG | 19.3±2.4 a | 101.3±6.4 a | 1284±158 a | 12.7±0.3 b | 62.1±3.3 a |
| AU | 12.0±2.1 ab | 37.7±9.3 b | 525±142 b | 13.9±0.7 a | 43.7±6.3 b |
| SAA | 10.7±0.8 b | 31.0±1.7 b | 332±52.1 b | 11.8±0.5 b | 31.1±4.0 b |
| Semillas | |||||
| Núm./racimo | Núm./rama | Núm./fruto | Núm./planta | g/planta | |
| MAG | 105.7±2.9 b | 553.8±52.9 a | 8.34±0.04 a | 10707±666 a | 109.96±0.7 a |
| AU | 133.2±7.2 a | 418.1±60.0 ab | 9.57±0.04 a | 5017±634 b | 51.67 ±0.6 b |
| SAA | 99.6±5.1 b | 286.3±37.3 b | 9.21±0.05 a | 3054±415 c | 31.45±0.6 c |
*MAG: con manejo agrícola y ganadero circundante, AU: con actividad urbana circundante y SAA: sin actividad antrópica circundante. Valores en la misma columna seguidos con la misma letra no muestran diferencia significativa (P ≤ 0.05).
Número de racimos
El número promedio de racimos por planta varió (P ≤ 0.05) entre las localidades. Las plantas de la localidad MAG presentaron en promedio el triple respecto a las otras localidades (Tabla 2). La distribución de los racimos en las ramas fue heterogénea entre las localidades. En las localidades MAG y AU (con actividad urbana aledaña), en promedio la proporción mayor de racimos se concentró en las primeras ramas basales; en contraste en las plantas de la localidad SAA (sin actividad antrópica aledaña) la distribución pareció relativamente homogénea. Además, el número promedio de racimos varió entre las ramas a lo largo del tallo; en las localidades MAG y AU hubo ramas con promedio mínimo (0.3) de racimos y máximo de nueve independientemente de la ubicación de la rama (Fig. 2 A, B y C).
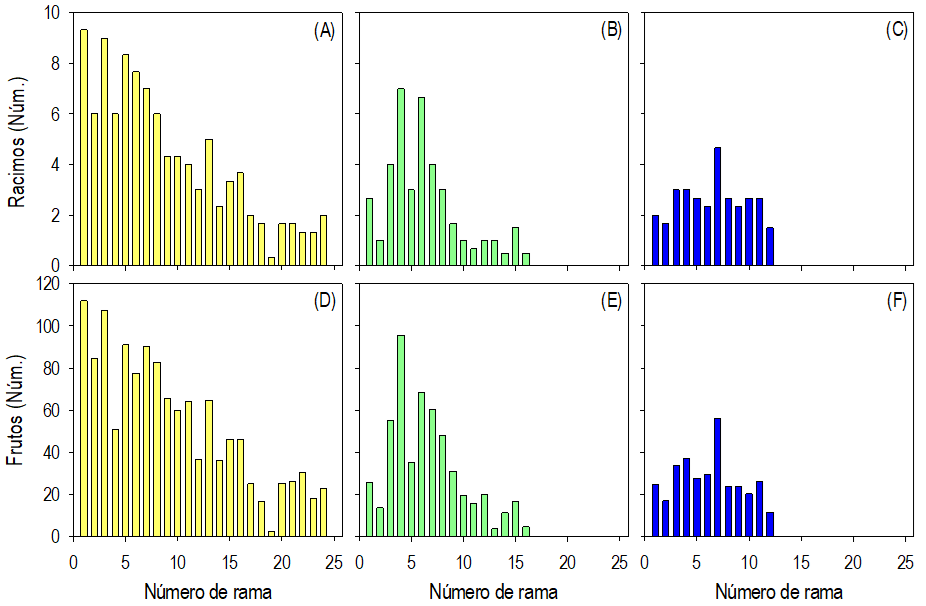
Fig. 2 Promedio del número y distribución de racimos frutales y de frutos en las ramas, en sentido acrópeto, sobre el tallo de plantas de Crotalaria longirostrata Hook & Arn. recolectadas en Guerrero, México. Localidades MAG: con manejo agrícola y ganadero circundante (A y D), AU: con actividad urbana circundante (B y E) y SAA: sin actividad antrópica circundante (C y F).
Número de frutos
El número promedio de frutos por planta, por racimo y por rama varió significativamente (P ≤ 0.05) entre las localidades. Las plantas de la localidad MAG presentaron en promedio entre el doble y el triple de frutos respecto a las otras localidades. En contraste los racimos de las plantas de la localidad AU tuvieron, en promedio, 12 % más frutos que en las localidades MAG y SA. Aunque el número de frutos por racimo fue mayor en las plantas de la localidad AU, el número promedio de frutos por rama fue 40 % mayor en la localidad MAG respecto a las otras localidades (Tabla 2).
La distribución de los frutos en las ramas a lo largo del tallo fue heterogénea en las plantas de las tres localidades y mostró tendencias similares a la distribución de los racimos (Fig. 2). En general, el número promedio de frutos en las ramas basales (en el primer tercio del tallo) de las plantas de las localidades MAG y AU, fue mayor al 50% del total. Algunas de esas ramas presentaron alrededor de 100 frutos; en contraste, algunas ramas del centro del tallo y las del tercio superior, hacia el ápice, tuvieron sólo dos o tres frutos. En localidad SAA la distribución de los frutos en las ramas pareció relativamente homogénea (Fig. 2 D, E y F).
Rendimiento de semillas
El número promedio de semillas por racimo, por rama y por planta varió significativamente (P ≤ 0.05) entre las localidades. En el caso de los racimos, los de las plantas en la localidad AU promediaron 23% más semillas que en las localidades MAG y SAA. En el caso de las ramas, las plantas de la localidad MAG presentaron en promedio entre 25 y 48% más semillas que en las localidades AU y SAA, respectivamente. El número promedio de semillas por planta mostró las diferencias mayores entre las localidades; en la localidad MAG se cuantificó un promedio de 10707 semillas por planta, valor que representó 53% más semillas que en las plantas de la localidad AU y 72% más que en la localidad SAA (Tabla 2).
El número promedio de semillas por fruto (8 a 10) fue el carácter más estable en las plantas de las tres localidades, pues las diferencias en este carácter no fueron significativas (P > 0.05; Tabla 2). Sin embargo, la frecuencia del número de semillas por fruto (Fig. 3) mostró que en la localidad MAG los frutos tuvieron desde una hasta 15 semillas, a diferencia de los frutos en las plantas de las localidades AU y SAA que contenían mínimos de 3 y 5 semillas, respectivamente y máximos de 14 en ambos casos. La distribución de la frecuencia del número de semillas por fruto fue cercanamente simétrica en las localidades AU y SAA, con el máximo de nueve semillas por fruto en ambas. La distribución en la localidad MAG contrastó con las anteriores, ya que mostró dos máximos, en 7 y 10 (Fig. 3).
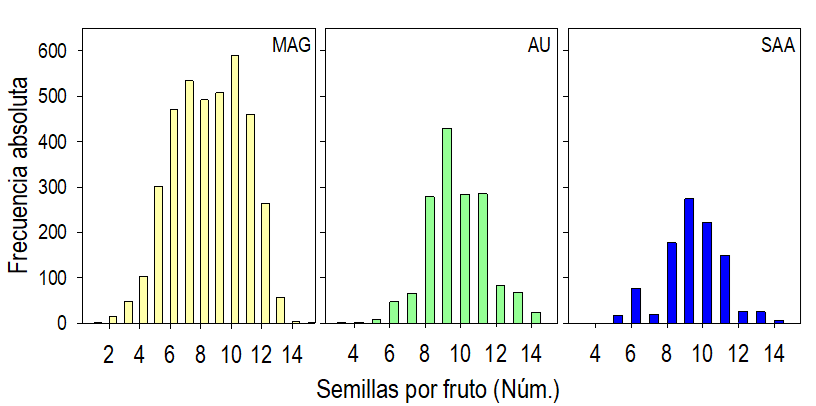
Fig. 3 Frecuencia del número de semillas por fruto en plantas de Crotalaria longirostrata Hook & Arn. recolectadas en Guerrero, México. Localidades MAG: con manejo agrícola y ganadero circundante, AU: con actividad urbana circundante y SAA: sin actividad antrópica circundante.
Las diferencias en los componentes del rendimiento entre las localidades, ya descritas, resultaron en el rendimiento significativamente diferente entre ellas. La localidad MAG mostró el rendimiento significativamente mayor; este representó 53 y 71% rendimiento mayor de semilla respecto a las localidades AU y SAA (Tabla 2).
La distribución de las semillas en las ramas a lo largo del tallo fue heterogénea en las plantas de las tres localidades (Fig. 4). Estas distribuciones mostraron tendencias similares a las descritas por los racimos y los frutos en las ramas en las tres localidades (Fig. 2). Las diferencias principales son los valores de cada estructura (máximos cercanos a 10 en el caso de los racimos, 115 en el de los frutos y 1030 en el de semillas por rama). Así, el número promedio de semillas en las ramas basales (alrededor del primer tercio del tallo) de las plantas de las localidades MAG y AU fue mayor al 50% del total. Varias de esas ramas presentaron más de 600 semillas y hasta cerca de 1000; al contrario, principalmente las ramas cercanas al ápice tuvieron los números menores de semillas. La distribución de las semillas en las ramas de las plantas de la localidad SAA difirió de las otras dos localidades en la relativa uniformidad; es decir, las ramas apicales acumularon cantidades de semilla cercanamente similares al resto (Fig. 4).
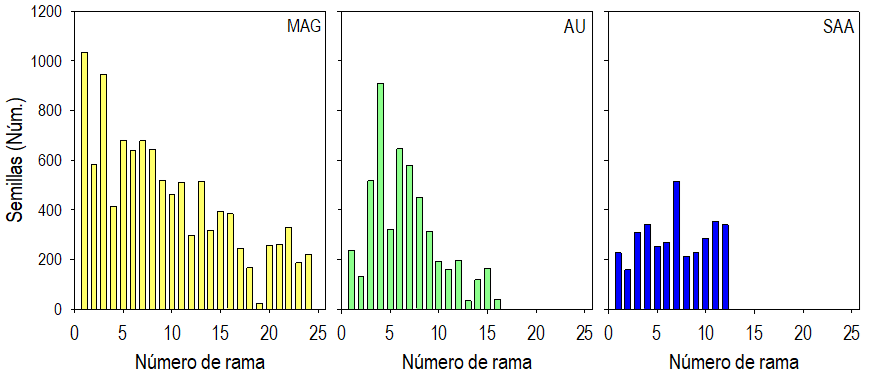
Fig. 4 Promedio del número y distribución de semillas en las ramas, en dirección acrópeta, sobre el tallo de plantas de Crotalaria longirostrata Hook & Arn. recolectadas en localidades de Guerrero, México. Localidades MAG: con manejo agrícola y ganadero circundante, AU: con actividad urbana circundante y SAA: sin actividad antrópica circundante.
Longitud, diámetro y biomasa de los frutos
Las dimensiones y la biomasa de los frutos, valvas y semillas las cuantificamos en los 3852 frutos de las plantas muestreadas en la localidad MAG. Los frutos mostraron color castaño obscuro, se caracterizaron por ser frutos inflados y tres veces más largos que anchos (Fig. 5A). El diámetro promedio (de 5.91 a 6.94 mm) y la longitud promedio (de 17.44 a 18.40 mm) difirieron (P ≤ 0.05) entre las plantas; sin embargo, independientemente de su origen silvestre las diferencias entre las dimensiones de esas estructuras son mínimas (Fig. 5A). La distribución de la frecuencia del diámetro y de la longitud de los frutos de la muestra completa (de 3852 frutos) fue simétrica alrededor de la media; mostró la variación del diámetro entre 4 y 8 mm, con la frecuencia absoluta máxima de entre 6.2 y 6.4 mm (Fig. 6A). En el caso de la longitud, la distribución fue de 12.5 a 22 mm, con frecuencia absoluta máxima entre 18 y 18.25 mm (Fig. 6B).
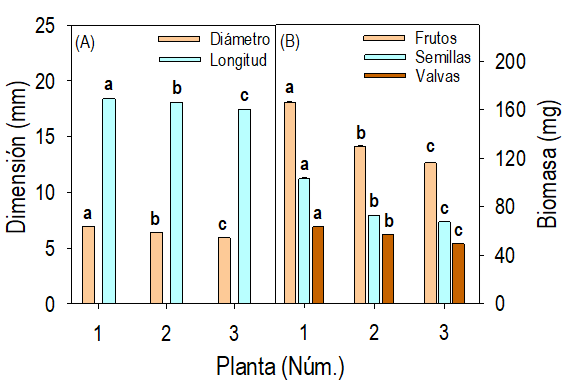
Fig. 5 Promedio (+ E.E.) de las dimensiones de los frutos (A), de su biomasa y la biomasa de sus semillas y valvas (B) por planta de Crotalaria longirostrata Hook & Arn., de la localidad Las Vigas (con manejo agrícola aledaño), Guerrero, México.
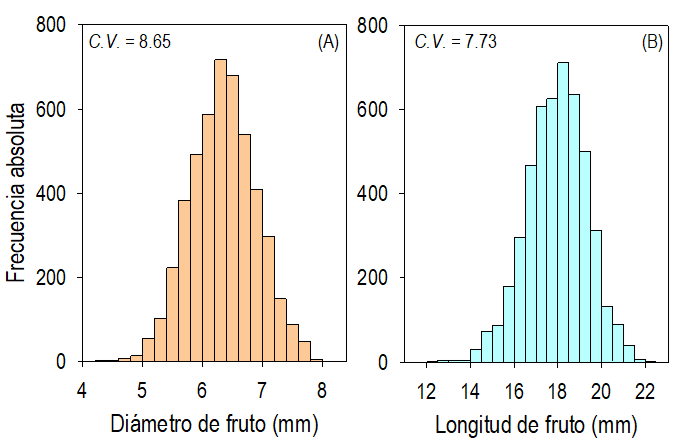
Fig. 6 Frecuencia del diámetro (A) y de la longitud (B) de los frutos de plantas de Crotalaria longirostrata Hook & Arn. recolectadas en Las Vigas, Guerrero, México (localidad con manejo agrícola aledaño).
Las diferencias (P ≤ 0.05) de la biomasa promedio de las valvas, los frutos, y las semillas representaron hasta 21.6, 29.9 y 34.4%, respectivamente entre las plantas (Fig. 5B). La distribución de la frecuencia de la biomasa de los frutos y de sus estructuras fue sesgada a la derecha (Fig. 7 A-C); aunque, el sesgo en el lado derecho fue producto de una muestra de menos de 10 frutos con peso atípico alto en la muestra de cerca de los 4000 evaluados. Así, la frecuencia absoluta máxima de la biomasa de los frutos, las semillas y las valvas se identificó en intervalos estrechos: 55-60 mg, 130-140 mg y 170-175 mg, respectivamente (Fig. 7 A-B).
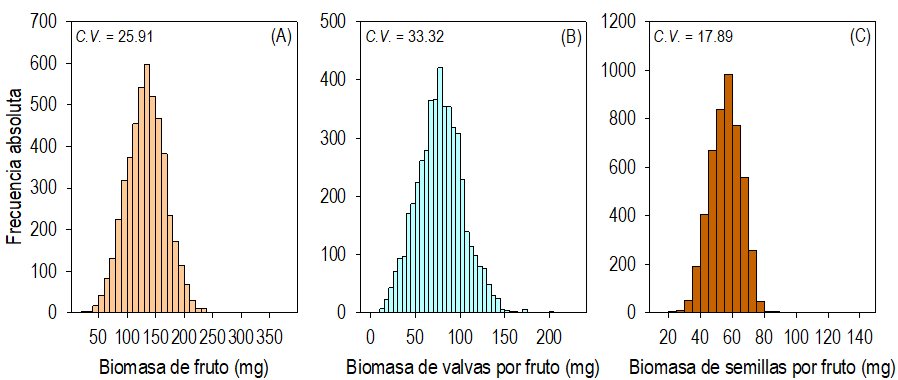
Fig. 7 Distribución de la frecuencia de la biomasa de los frutos (A), de sus valvas (B) y semillas (C) en las ramas a lo largo del tallo, en dirección acrópeta, de plantas de Crotalaria longirostrata Hook & Arn. recolectadas en Las Vigas, Guerrero, México (localidad con manejo agrícola aledaño).
Las dimensiones y la biomasa de los frutos y de sus estructuras se correlacionaron positivamente y en forma altamente significativa unas con otras (Tabla 3); lo que mostró la alta asociación entre el crecimiento de los frutos y el rendimiento de semilla.
Tabla 3 Coeficientes de correlación de Pearson entre las dimensiones y la biomasa de los frutos de plantas de Crotalaria longirostrata Hook & Arn. recolectadas en Las Vigas, Guerrero, México (localidad con manejo agrícola aledaño).
| Variable | Diámetro | Longitud | Biomasa de fruto |
Biomasa de semillas |
Biomasa de valvas |
|---|---|---|---|---|---|
| Diámetro | 1.000 | 0.606*** | 0.772*** | 0.697*** | 0.797*** |
| Longitud | 1.000 | 0.643*** | 0.571*** | 0.698*** | |
| Biomasa de fruto | 1.000 | 0.962*** | 0.875*** | ||
| Biomasa de semillas | 1.000 | 0.765*** | |||
| Biomasa de valvas | 1.000 |
*** P < 0.0001
Análisis de componentes principales
El análisis de CP incluyó las variables con correlación no significativa; ya que, si las variables están correlacionadas significativamente entre ellas, la información que aportarán será redundante y la condición de no colinealidad se violará para la regresión por mínimos cuadrados. En el análisis de CP, los dos primeros CP acumularon el 84.8% de la varianza total reconocida. El CP1 se definió principalmente por el número de racimos por rama, frutos por rama y semillas por rama. El CP2 reveló la dependencia directa del número de semillas por racimo con el número de frutos por racimo (Tabla 4). La representación gráfica del CP2 en función de CP1 mostró que cada una de las variables evaluadas tiene efectos equivalentes en eso CP. La localidad AU traslapó completamente a la localidad SAA y gran parte de MAG; aunque, los componentes del rendimiento definieron a cada localidad (Fig. 8).
Tabla 4 Valores propios y proporción de la varianza para los primeros componentes principales generados a partir de los componentes del rendimiento de plantas de Crotalaria longirostrata Hook & Arn. recolectadas en tres localidades de Guerrero, México.
| Componente principal | ||
|---|---|---|
| 1 | 2 | |
| Racimos por rama (Núm.) | 0.529 | 0.215 |
| Frutos por rama (Núm.) | 0.574 | 0.034 |
| Frutos por racimo (Núm.) | 0.181 | -0.616 |
| Semilla por rama (Núm.) | 0.562 | -0.024 |
| Semilla por racimo (Núm.) | 0.103 | -0.676 |
| Semilla por fruto (Núm.) | -0.180 | -0.340 |
| Varianza acumulada | 0.498 | 0.848 |
Los valores con trazo grueso en los componentes principales tuvieron aporte mayor.
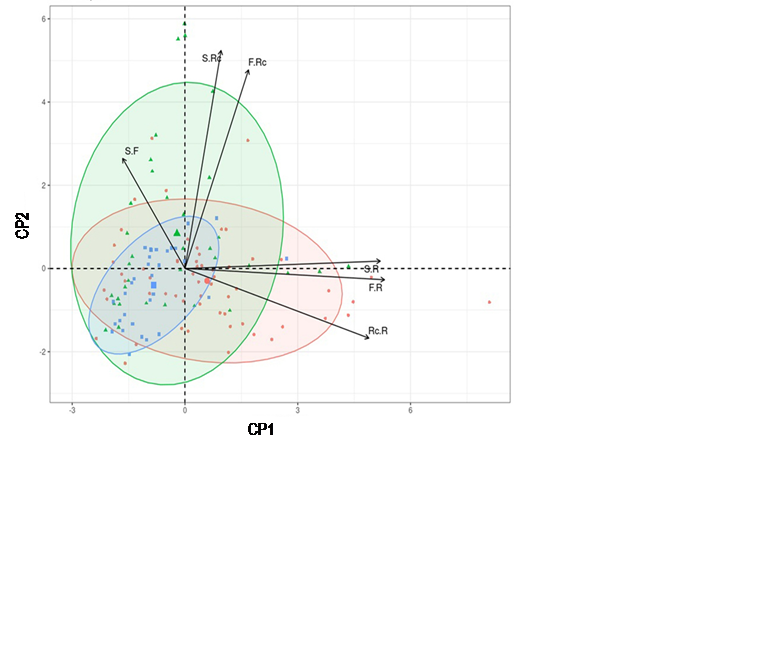
Fig. 8 Ordenación de los componentes principales (PC) 1 y 2 de plantas de Crotalaria longirostrata Hook & Arn. recolectadas en tres localidades de Guerrero, México, según los componentes del rendimiento (Círculos rojos y área dentro de la elipse roja, tríangulos verdes y área dentro de la elipse verde y cuadros azules y área dentro de la elipse azul corresponden respectivamente a las localidades Las Vigas, Ometepec y Llano Grande de los Hilarios Igualapa, Guerrero, México; Rc. R: racimos por rama, F. R: frutos por rama; F. Rc: frutos por racimo; S. R: semillas por rama; S. Rc: semillas por racimo; S. F: semillas por fruto).
Características del suelo
Entre las diferencias en el suelo de las tres localidades destacaron las siguientes. Los suelos se clasificaron de acuerdo con su textura como franco arenoso en las localidades MAG y AU y como franco limoso en la localidad SAA. La proporción de arcilla no difirió (P > 0.05) entre las localidades, la proporción de arenas fue 24% mayor (P ≤ 0.05) en MAG y AU, respecto a SAA; en contraste, en esta última los limos fueron 34.29% más abundantes que en las otras dos. Las localidades MAG y SAA fueron en promedio 21% más ricas (P ≤ 0.05) en materia orgánica respecto a AU; además, los suelos difirieron en las concentraciones de los macroelementos N, P y K. La localidad AU presentó 55% más N total y, junto con SAA, 9% menos K soluble. Aunque, el K intercambiable no difirió entre las localidades. La concentración de P extraíble mostró el gradiente: MAG > AU > SAA, en el que la última localidad mostró hasta cinco veces menos P que MAG. La densidad aparente y la densidad real, relacionada con la compactación del suelo, fueron mayores en el suelo de la localidad AU. Además, la capacidad de campo y el punto de marchitez permanente del suelo mostraron el siguiente gradiente entre las localidades: SAA > AU > MAG, en el que los valores de estas dos características en la localidad SAA fueron 31.06 y 27.29% mayores que en MAG. Aunque el pH y el Mg intercambiable no fue diferente entre las localidades, el Ca intercambiable mostró el siguiente gradiente AU > MAG > SAA, en el que los valores en AU fueron 38% mayores que en SAA (Tabla 5).
Tabla 5 Características físicas y químicas del suelo de tres localidades de Guerrero, México.
| Localidad* | |||
|---|---|---|---|
| MAG | AU | SAA | |
| Textura | Franco arenoso | Franco arenoso | Franco limoso |
| Color (seco) | |||
| Matiz (hue) | 10YR | 10YR | 10YR |
| Brillo (value) | 4 | 6 | 4 |
| Pureza | 3 | 7 | 5 |
| Composición | |||
| Arcilla (%) | 7 a | 8 a | 9 a |
| Arenas (%) | 59 a | 56 a | 44 b |
| Limos (%) | 34 b | 36 b | 47 a |
| Materia orgánica (%) | 2.017 a | 1.748 b | 2.420 a |
| N total (%) | 0.023 b | 0.018 b | 0.046 a |
| P extraíble (mg kg-1) | 0.020 a | 0.012 b | 0.004 c |
| K soluble (cmol(+) kg-1 ) | 0.271 a | 0.165 b | 0.163 b |
| Densidad, Estado hídrico y Conductividad | |||
| Aparente (g/cm3) | 1.52 b | 1.91 a | 1.49 b |
| Real (g/cm3) | 2.68 b | 2.85 a | 2.78 ab |
| Capacidad de campo (%) | 18.84 c | 21.33 b | 27.33 a |
| Punto de marchitez permanente (%) | 9.43 c | 10.39 b | 12.97 a |
| Conductividad eléctrica (dSm-1) | 0.02 b | 0.02 b | 0.07 a |
| Capacidad de intercambio catiónico (cmol(+) kg-1) | 2.90 a | 2.85 a | 2.20 b |
| pH (1:2 suelo:agua) | 5.46 a | 5.23 a | 5.10 a |
| Bases intercambiables | |||
| Ca (cmol(+) kg-1) | 1.59 b | 1.92 a | 1.19 c |
| Mg (cmol(+) kg-1) | 0.72 a | 0.53 a | 0.56 a |
| K intercambiable (cmol(+) kg-1) | 0.325 a | 0.266 a | 0.284 a |
*MAG: con manejo agrícola y ganadero circundante, AU: con actividad urbana circundante y SAA: sin actividad antrópica circundante. Letras iguales en una línea denotan que no hay diferencia significativa (P ≤ 0.05).
Discusión
Las plantas recolectadas fueron arbustiformes, tuvieron tallo central erecto, leñoso sólo en su parte inferior (tallo sufruticoso), con altura mayor a 150 cm, frutos aun indehiscentes y valvas color castaño oscuro, similares a las descritas por Azurdia (2016) y Soto-Estada (2004). Entre las localidades se observaron diferencias (P ≤ 0.05) en 90 % de las 10 variables del rendimiento evaluadas. Esta variabilidad se explica en parte por el carácter silvestre de las plantas (Peña-Valdivia et al., 2012). Además, el comportamiento fisiológico diferenciado, que conduce a contrastes en los componentes del rendimiento de las variantes o biotipos (plantas de cada localidad) depende de las diferencias en los ambientes de crecimiento, lo que se conoce como interacción genotipo-ambiente (Mederos-Ramírez & Ortiz-Pérez 2021).
Entre los factores ambientales con mayor efecto en el crecimiento de las plantas, y por tanto en los componentes del rendimiento y rendimiento de semilla, están la precipitación y la temperatura ambiente. De acuerdo con los datos de CONAGUA (2021), las localidades MAG y AU muestran precipitación promedio anual total (1310 y 1350 mm, respectivamente) y distribución de la precipitación durante el año similares; además, a pesar de ciertas fluctuaciones de la temperatura media (26.5 y 27 ºC) y máxima (32.8 y 33.1 ºC) durante el año sus promedios son similares (Fig. 9 A-B). Además, estas dos localidades comparten el mismo tipo de clima (Tabla 1). En contraste, con estas similitudes de su ambiente, las plantas de ambas localidades difirieron significativamente en 60% los componentes del rendimiento evaluados y en el rendimiento de semilla (Tabla 2). Además de lo anterior, el clima de la localidad SAA es diferente al de las otras dos, muestra mayor precipitación (total 2063 mm) y su ambiente es menos cálido (Fig. 9C); pero, 60% de los componentes evaluados fueron similares en los de la localidad AU (Tabla 2). Lo anterior puede ser evidencia de la presencia de tres biotipos, parcialmente distintos, con interacción específica con su ambiente. Respuestas equivalentes se han documentado en biotipos de frijol silvestre (Peña-Valdivia et al., 2012).
La similitud de las dimensiones de los tallos entre las localidades es relevante por su relación con el rendimiento de los frutos y las semillas. Al respecto, los tallos desempeñan un papel de apoyo en el desarrollo de los órganos reproductores, como conductos para el transporte de recursos y como estructuras físicas que soportan el peso de los frutos en desarrollo. En leguminosas, como chícharo (Pisum sativum) y soya (Glycine max (L.) Merr.), el tamaño del tallo (altura y diámetro) es relevante, ya que puede recibir hasta un tercio de los fotoasimilados producidos por las hojas durante el desarrollo de los frutos y competir por ellos (Alerding et al., 2017). Así, el crecimiento, axial y radial, y su conclusión antes o durante la etapa reproductiva tiene distintos efectos en el rendimiento de frutos y semillas (Purcell et al., 2014). Si los tallos continúan la expansión radial durante la reproducción, los recursos se asignarán a la construcción de la pared celular secundaria, que incluye las síntesis de lignina, y al llenado de semillas. En especies de tallo erecto, que producen anillos concéntricos de tejidos secundarios, el crecimiento axial del tallo termina cuando la planta inicia la etapa de llenado de semillas (Purcell et al., 2014). Actualmente, los aspectos relacionados con la terminación del crecimiento radial dentro del sistema de tallo y ramas en C. longirostrata se desconoce; por lo que, en estudios futuros podría evaluarse la anatomía y fisiología del desarrollo de esas estructuras para complementar el conocimiento de su aporte al rendimiento de frutos y semillas.
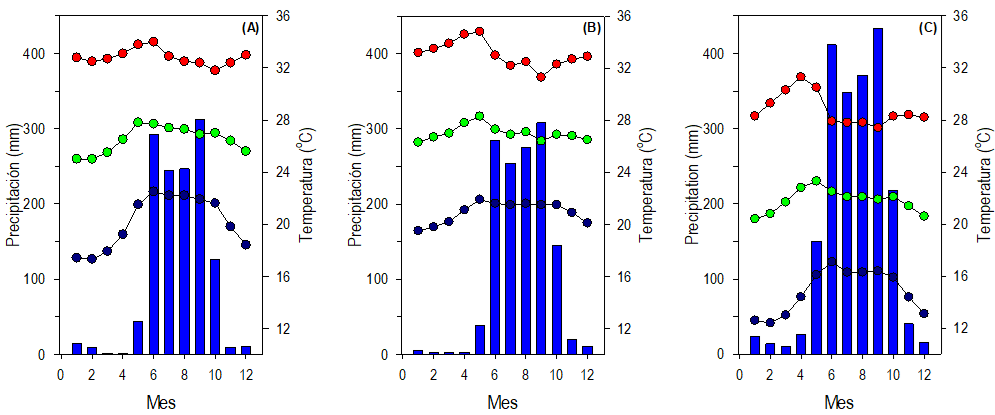
Fig. 9 Climogramas basados en precipitación promedio (barras) y temperatura promedio mínima (círculos negros), media (círculos verdes) y máxima (círculos rojos) en Las Vigas (A), Ometepec (B) y Llano Grande de los Hilarios Igualapa (C), Guerrero, México. Cada barra y círculo representa los datos promedio mensuales (1: enero… 12: diciembre) durante un período de seis décadas, de 1950 a 2010 (CONAGUA, 2021).
El número significativamente mayor de 38 y 45% de ramas en las plantas de la localidad MAG respecto a AU y SAA no fue proporcional a las diferencias en el número de racimos o frutos por planta entre las localidades; ya que, en la localidad MAG la cantidad mayor de racimos representó 63 y 70% respecto a AU y SAA y las diferencias en la cantidad de frutos alcanzó 60 y 74%, respectivamente. Estos resultados indican que la exposición de ramas, como uno de los componentes del rendimiento, repercute en el desarrollo de estructuras reproductivas en C. longirostrata. Al respecto, el espaciamiento entre las plantas es uno de los factores que afectan la ramificación y el rendimiento de las semillas en especies como C. juncea y soya (Tripathi et al., 2013; Xu et al., 2021), por su efecto en la tasa de crecimiento y en la competencia entre las plantas por los insumos necesarios para el crecimiento y el desarrollo. Xu et al. (2021) analizaron la contribución de las ramas al rendimiento de soya mediante dos fechas de siembra en seis densidades de siembra de dos cultivares; el número de vainas por planta creció con el aumento del número de ramas, mientras que el número de vainas por unidad de área disminuyó, por lo que, el rendimiento también se redujo. Además, los autores observaron respuesta diferente de los cultivares; uno, con una densidad menor de plantación, incrementó el rendimiento como resultado de más ramas; otro cv. incrementó su potencial de rendimiento con una mayor densidad de plantación debido al menor número de ramas y su tolerancia mayor a la competencia por densidad de siembra mayor. Así, el rendimiento de semillas puede aumentarse en plantas con poca capacidad de ramificación en siembra con densidad moderada (Xu et al., 2021). La relevancia de las ramas en el incremento del rendimiento de semilla en C. juncea se debió a que el desarrollo de las ramas laterales aumentó los sitios para el desarrollo de frutos; esto se comprobó cuando se eliminó la dominancia apical, al separar el ápice del tallo (Tripathi et al., 2013). Por lo que, en estudios futuros la densidad de plantas de C. longirostrata en los sitios muestreados complementará el análisis de las diferencias en el rendimiento de semilla.
Los frutos del presente estudio tuvieron longitud y diámetro que coincidieron con los de ejemplares de herbario (longitud entre 1.5 y 2.5 cm y diámetro entre 5 y 9 mm) recolectados en el Valle de Tehuacán-Cuicatlán, Puebla-Oaxaca, México (Soto-Estada, 2004) y en la región del Bajío, México (Rzedowski et al., 2016). Estos resultados indican que las dimensiones de los frutos de C. longirostrata son caracteres relativamente estables y típicos en diversas regiones de México. Esta homogeneidad contrasta con la distribución de los racimos y frutos a lo largo del tallo en las localidades muestreadas. En relación con los frutos, las valvas son componentes del rendimiento que pueden tener efecto en el rendimiento de semilla. Esas estructuras, además de la función protectora a las semillas, pueden tener actividad fotosintética y aportar asimilados y nutrientes que impulsan el crecimiento de las semillas; además, la señalización generada por las valvas coordina el crecimiento, llenado y maduración de las semillas, todo esto con repercusiones en el rendimiento (Bennett et al., 2011).
El número de semillas por fruto fue el componente del rendimiento que no difirió entre las localidades. Pero las diferencias en la cantidad de racimos y frutos por rama y por planta entre las localidades pueden repercutir en el rendimiento de semilla; aunque, con tendencias parcialmente diferentes. Esto permite suponer la presencia de tres biotipos, uno por cada localidad muestreada, que de acuerdo con el ACP no difieren completamente, ya que comparten ciertas características en los componentes del rendimiento. En contraste con la homogeneidad de las dimensiones de los frutos, el número de semillas, nueve en promedio, por fruto en las tres localidades fue poco menor al documentado, de entre 10 y 16, en algunos ejemplares de herbario (Soto-Estada, 2004; Rzedowski et al., 2016).
La cantidad de frutos por planta, como componente del rendimiento, está relacionada directamente con la producción de semillas (Guerrero, 1999). Estas últimas tienen importancia central en el presente estudio, ya que es el insumo crítico en los sistemas agrícolas. Es decir, para el cultivo y multiplicación de plantas, en campo de cultivo o invernadero, es necesario contar con semilla suficiente y de buena calidad (Vinicius de Araujo et al., 2018). En este estudio se demostró que las poblaciones silvestres de C. longirostrata, del estado de Guerrero, producen en promedio de 3054 a 10707 semillas por planta. En contraste, el número promedio de semillas por fruto es un carácter homogéneo (8 a 10 semillas por fruto) entre las localidades. Además, la calidad fisiológica de las semillas de las tres regiones muestreadas está actualmente en proceso, para complementar la información.
Los resultados del presente estudio indican que las plantas más productivas, con mayor número de ramas, racimos y frutos por planta, número de frutos por racimo y por rama, número de semillas por rama y por planta y rendimiento de semilla se desarrollaron en la localidad MAG, con suelo franco arenoso, con capacidad de campo y punto de marchitez permanente menores, pero con capacidad de intercambio catiónico, y contenido de P y K intercambiable mayores respecto a los otros sitios. Además, el sitio MAG es una zona con potreros y cultivos, como maíz y frijol, cercanos. En contraste, las plantas con los valores significativamente menores de esos componentes del rendimiento y del rendimiento correspondieron a la localidad SAA, en suelo franco limoso, con pendiente de 25°, entre cerros y con capacidad de campo, punto de marchitez permanente, conductividad eléctrica y N total mayores respecto al de los otros sitios; además, esas plantas con los rendimientos menores crecieron en suelo con menor contenido de Ca intercambiable y P extraíble, respecto a los otros sitios. Al contrario, el pH y el contenido de Mg y de K intercambiable fueron similares en las tres localidades, por lo que no se relacionaron directamente con las diferencias en el rendimiento y sus componentes entre las localidades.
Conclusiones
Los componentes del rendimiento y el rendimiento de semilla de poblaciones de C. longirostrata muestran variabilidad genética (biotipos) y especificidad al ambiente de crecimiento. Los biotipos, independientemente del ambiente en el que crecen, producen miles de semillas por planta con potencial de uso para multiplicar las plantas ex situ. Sin embargo, los ambientes menos cálidos y los suelos con capacidad de campo y punto de marchitez permanente mayores parecen menos favorables para la producción de semillas. Al contrario, el suelo con mayor capacidad de intercambio catiónico, concentración de materia orgánica, P extraíble y K soluble parece favorecer el desarrollo de las plantas de C. longirostrata, que resulta en alto rendimiento de semilla.
Actualmente, la información del rendimiento y sus componentes del germoplasma de C. longirostrata es escasa; por lo tanto, los resultados del presente estudio son la información más completa con la que se cuenta a la fecha y puede ser la base para la producción agrícola de la especie. Aunque, es necesaria la evaluación de la calidad fisiológica de la semilla y su respuesta a las variaciones ambientales anuales.

