











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
The tropical deciduous forest (TDF) occurs in 25 of the 32 Mexican states. It represents 80% of the vegetation cover on the Pacific slope, part of central Mexico, the Balsas Depression and the Tehuacán-Cuicatlán Valley. Besides, the remaining 20% of the TDF occurs on the slope of the Gulf of Mexico (Banda et al., 2016; Challenger & Soberón, 2008; Dirzo, 1994; Meave et al., 2012; Miranda & Hernández, 1963; Pennington & Sarukhán, 2005; Rzedowski, 1978; Rzedowski & Calderón-de Rzedowski, 1987, 2013; Trejo, 1998, 1999).
Research related to the dynamics, functioning and ecology of TDFs has allowed us to generate knowledge about this plant community, which grows on mountains, hillsides and gradients with moderate to strong slopes (Challenger & Soberón, 2008; Rzedowski, 1978; Trejo, 1996, 2010). Climate is mainly warm subhumid (Awo) according to the Köppen classification, modified by García (2004). The Burseraceae and Fabaceae families are well-represented in the TDF, along with Anacardiaceae, Cactaceae, Asteraceae, Euphorbiaceae, Malpighiaceae and Rubiaceae, among others (Miranda & Hernández, 1963; Rzedowski, 1978; Trejo, 1996, 2005, 2010).
A total of 749 genera and 2,500 species have been recorded, included in the primary and secondary vegetation of this ecosystem. The distinctive feature of the TDF is its marked seasonality reflected in the phenology of the plants, with the presence of a long dry season lasting 3 to 8 months (over 75%), while the rainy season lasts 4 to 6 months. The physiognomy of the TDF is characterised by trees that vary in height from 8 to 12 m on average (Banda et al., 2016; Challenger & Soberón, 2008; Miranda & Hernández, 1963; Pennington & Sarukhán, 2005; Rzedowski, 1978; Rzedowski & Calderón-de Rzedowski, 2013; Trejo, 1996, 2005, 2010).
It should be noted that in the last 2 decades, there has been an increasing number of regional- or state-level studies on the functioning of the TDF in the context of physiognomy, structure, floristic composition and diversity. Some notable studies, grouped by their geographical distribution, are as follows: 1) Pacific Slope: in Sonora (Sánchez-Mejía et al., 2007); Banderas Bay, Nayarit (Bravo-Bolaños et al., 2016); Baja California Peninsula (León-de la Luz et al., 2012); Oaxaca (Gallardo-Cruz et al., 2005) and the Central Depression of Chiapas (Rocha-Loredo et al., 2010); 2) Central Mexico: at the Balsas Depression (Méndez-Toribio et al., 2014; Pineda-García et al., 2007); Puebla (Rodríguez-Acosta et al., 2014); Sierra de Huautla Biosphere Reserve, Morelos (Beltrán-Rodríguez et al., 2018) and 3) Gulf Slope: at the Dzilam Reserve, Yucatán (Leirana-Alcocer et al., 2009), Campeche (Dzib-Castillo et al., 2014), and Veracruz (Williams-Linera & Lorea, 2009).
The analysis of the area and percentage of the tropical deciduous forest (TDF) in the country shows a clear decrease in its distribution area. The first estimate was calculated at 31,521,290 ha (15.98%) by Flores et al. (1971). Other reports are those of Rzedowski and Calderón-de Rzedowski (2013) with 28,000,000 ha (14%); INEGI (1980) with 15,980,000 ha (8.2%); Trejo (2010)ca. 6,850,000 ha (30%); Conafor-Semarnat (2012) with 12% (15,869,741.80 ha). Trejo and Dirzo (2000) reported that 3.7% (72,900 km2) corresponds to primary vegetation. Finally, the data provided by INEGI (2003) is 33.5 million hectares, while Challenger and Soberón (2008) reported an estimate of 11.26% (7.93 and 14.19 million hectares as primary and secondary vegetation, respectively) of the surface area of the entire Mexican territory. Recently, INEGI (2014) reported that the TDF occupies 11.9% of Mexico’s vegetation cover.
Among the processes that have impacted the functioning and fragility of this ecosystem are exploitation of forest resources, land-use changes, habitat fragmentation, introduction of exotic species, forest fires, urban, industrial and tourism development and communication routes, among others (Dirzo et al., 2011; Hansen et al., 2013; Maass et al., 2010; Ramírez-Bravo & Hernández-Santin, 2016; Rzedowski, 1978; Rzedowski & Calderón-de Rzedowski, 2013). It is also important to highlight the limited resilience of the secondary community of the TDF compared to a climax community (Rzedowski & Calderón-de Rzedowski, 2013). Therefore, it is considered the most threatened tropical ecosystem worldwide (Banda et al., 2016; Beltrán-Rodríguez et al., 2018; Ceballos & García, 1995; Janzen, 1988; Masera et al., 1997; Miles et al., 2006; Trejo, 2005; Trejo & Dirzo, 2000).
In the state of Puebla, TDF occupies 15.7% (536,851 ha) (Rodríguez-Acosta et al., 2014) and represents 29% of the Tehuacán-Cuicatlán Valley area, where there are also various types of vegetation and a high levels of endemism, including TDF (Arriaga et al., 2000; Dávila et al., 1993, 2002; Osorio Beristain et al., 1996). Previous studies have been conducted in this region throughout the past seventy years, including floristic and phytogeographic ones (Jaramillo-Luque & González-Medrano, 1983; Miranda, 1948; Smith, 1965; Valiente-Banuet et al., 2000, 2009; Villaseñor et al., 1990; Zavala-Hurtado, 1982).
The objective of this study was to determine, the structure, diversity and floristic composition of the tree and shrub strata in the tropical deciduous forest of El Picante hill, located in the town of San José Tilapa, Puebla, in the south-eastern Tehuacán Valley, Mexico. This knowledge will not only complement the understanding of the dynamics and functioning of this ecosystem in relation to other regions of Mexico, but it will also be useful for decision-making regarding its conservation.
Material and methods
The study was conducted in the tropical deciduous forest (TDF) of El Picante hill, located 3 km northwest of the dirt road to San Antonio Barranca Vigas, between coordinates 18°10’49” and 18°10’59” N, 97°04’44.8” and 97°04’50.9” W, with an altitude range between 1,033 and 1,099 m asl, in the town of San José Tilapa, municipality of Coxcatlán, Puebla, Mexico, in the south-eastern Tehuacán Valley (Fig. 1a-d). The municipality of Coxcatlán is located in the province of the Sierra Madre del Sur, which includes the subprovinces of the eastern and central Sierras of Oaxaca. The temperature ranges from 14 to 26 °C, and the annual rainfall fluctuates from 300 to 1,100 mm. The climate can be very warm arid and warm arid (48% of surface area), semi-warm semi-arid (28%), temperate subhumid with summer rains (14%) and semi-warm sub-humid with summer rains (10%). The dominant soil type is Regosol, but Leptosol and Cambisol are also found. The types of vegetation present are tropical deciduous forest (32%), scrub (20%), forest (9%) and grasslands (3%) (Conabio, 2011; INEGI, 2009).
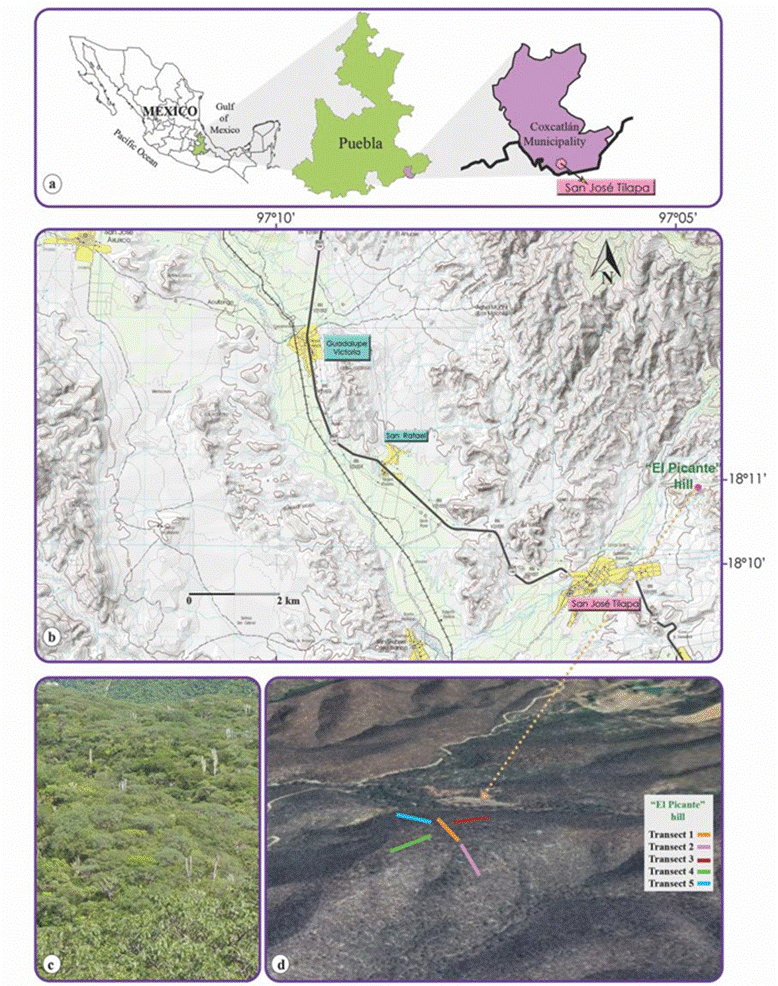
Figure 1 Geographic location of the study area, in San José Tilapa, in the municipality of Coxcatlán, Puebla, Mexico (a, b), tropical deciduous forest on El Picante hill (c), and satellite image with the 5 transects established in the study area (d).
Trees and shrubs were recorded during an annual cycle. Botanical samples were collected for latter identification. Since transects and plots have generally been used for studies of TDF vegetation in Mexico (Dzib-Castillo et al., 2014; León-de la Luz et al., 2012; Méndez-Toribio et al., 2014; Pineda-García et al., 2007; Rocha-Loredo et al., 2010), in the present work, a total of 5 transects (100 m long each) were established in different positions; at the summit (T1), in the north-eastern (T3), north-western (T5), western (T4) and south-eastern (T2) slopes of the hill (Fig. 1d), following the “point-centred quarter” sampling method, according to Matteucci and Colma (1982) and Mueller-Dombois and Ellenberg (1974). According to the method all transects (T1-T5) were divided every 10 meters and a total of 11 points per transect were considered. At each division, 4 quadrats were formed by one imaginary line crossed perpendicularly. In each quadrant, a tree and a shrub closest to the central point were located and the distance of them to the central point was recorded. For each tree and shrub (8 in total) the following measurements were taken: height, small (c) and large (C) diameters of canopy cover. Also, diameter at breast height (DBH at 1.30 m)for trees was obtained (Matteucci & Colma, 1982). To others short trees, shrubs, arborescent and rosette plants only density was recorded.
In addition, geographical coordinates, elevation and slope were recorded for each of the 10 central points of the transects. Collection of botanical samples was performed as completely as possible, with flowers and/or fruits and their respective duplicates, which were dried and pressed according to the methods proposed by Lot and Chiang (1986) for subsequent taxonomic determination, mounting and labelling. It should be noted that the floristic inventory was extended because of additional botanical collection performed in the transects studied. Plant identification was performed using various taxonomic keys included in several regional floras and in specialised literature. Once identified, the specimens were mounted and registered for incorporation into the Herbario Metropolitano (UAMIZ) from the Universidad Autónoma Metropolitana-Iztapalapa (UAM-I).
Vertical structure was analysed using the data from the height measurements of the trees and shrubs and the location of this vegetation in the strata. Tree strata were classified by height as follows: short: 2-4 m, medium: 4-6 m, and tall: ≥ 6 m. Shrub strata were similarly grouped: short: ≤ 1 m, medium: > 1-2 m, and tall: 2-4 m according to the González-Medrano (2003) classification. Horizontal structure was calculated using the following parameters: density, dominance and frequency, along with their respective relative values, in order to obtain the importance value index (IVI) of each of the species present at each sampling site. The following equations modified from Gaillard-de Benítez and Pece (2011) were used in order to calculate density.
For the calculation of the mean distance (D) of the trees and shrubs per hectare, the distances of each individual of each species from the central point were used; the sum of these distances was divided by the total number of individuals, as follows: the mean distance (D) of the trees and shrubs was calculated as follows:
where: di = the distance from i species to the centre of the transect; N = the total number of individuals.
The mean distance (D) values obtained were used to calculate the total tree and shrub density per hectare (DenTha) as follows:
Schematic diagrams were made for each of the transects using Adobe Illustrator software.
Richness (N) along with the Shannon-Wiener diversity index (H’), Pielou’s evenness index (J’), and the Jaccard similarity index (Magurran, 1988, 2004) were calculated using Multi-Variate Statistical Package (MVSP) software v. 3.2.
Results
General floristic composition
The plant composition of the tropical deciduous forest (TDF) on El Picante hill is made up of 53 taxa belonging to 47 genera and 28 families organized according to the APG III system (APG 2009; Table 1, Fig. 2). The Fabaceae family was the best represented with 10 taxa, contributing 18.9% of the flora, followed by Cactaceae (7.5%) with 4 taxa, Asteraceae (5.6%), and Burseraceae (5.6%), with 3 taxa each, followed by 9 families with 2 taxa each (Σ = 34%), and finally 15 families with a single taxon each (Σ = 28.3%). It is important to mention that 14 species endemic to the country were recorded in this study. They belong to 9 families: Cactaceae and Fabaceae were represented by 3 species each, Burseraceae with 2 species and 6 other families contributed 1 species each (Table 1).
Table 1 Floristic composition of the tropical deciduous forest (TDF) on El Picante hill in San José Tilapa, Puebla, Mexico. The inventory includes families, genera, species, common name and biological form. Species endemic to Mexico are indicated with an Asterisk (*) and taxa for the analysis of vegetation structure are marked with a black dot (•) and in bold type.
| Families/species | Common name | Biological form |
| Acanthaceae | ||
| • Justicia candicans (Nees) | shrub | |
| L.D. Benson | ||
| ANACARDIACEAE | ||
| *• Cyrtocarpa procera Kunth | “chupandillo” | tree |
| Apocynaceae | ||
| Asclepias curassavica L. | suffrutex | |
| • Plumeria rubra L. | “flor de mayo” | tree |
| Asparagaceae | ||
| Agave sp. | rosette | |
| Echeandia sp. | herb | |
| Asteraceae | ||
| *• Parthenium fruticosum | shrub | |
| Less. | herb | |
| Sanvitalia fruticosa Hemsl. | herb | |
| Zinnia peruviana (L.) L. | ||
| Bignoniaceae | ||
| Astianthus viminalis Baill. | tree | |
| Bromeliaceae | ||
| *Tillandsia circinnatioides | epiphyte | |
| Matuda | epiphyte | |
| Tillandsia recurvata (L.) L. | ||
| Burseraceae | ||
| *• Bursera aptera Ramírez | “cuajiote | tree |
| • Bursera morelensis | amarillo” | tree |
| Ramírez | tree | |
| *• Bursera submoniliformis | ||
| Engl. | ||
| Cactaceae | ||
| *Coryphantha pallida Britton | globular | |
| & Rose | columnar- | |
| *Myrtillocactus geometrizans | branched | |
| (Mart. ex Pfeiff.) | arborescent | |
| Console | columnar- | |
| Opuntia sp. | branched | |
| *Pachycereus weberi | ||
| (J.M.Coult.) Backeb. | ||
| Capparaceae | ||
| Quadrella incana (Kunth) | “mata | tree |
| Iltis & Cornejo | gallina” | |
| Commelinaceae | ||
| Commelina sp. | herb | |
| Tradescantia sp. | herb | |
| Convolvulaceae | ||
| Jacquemontia smithii | shrub | |
| B.L.Rob., & Greenm. | ||
| Cordiaceae | ||
| • Cordia curassavica (Jacq.) | shrub | |
| Roem., & Schult. | shrub | |
| Cordia stellata Greenm. | ||
| Ehretiaceae | ||
| Bourreria obovata Eastw. | shrub | |
| Euphorbiaceae | ||
| • Croton morifolius Willd. | shrub | |
| • Euphorbia schlechtendalii | tree | |
| Boiss. | ||
| Fouquieriaceae | ||
| *• Fouquieria formosa | tree | |
| Kunth | ||
| Fabaceae | ||
| • Acacia cochliacantha | tree | |
| Humb., & Bonpl. ex Willd. | shrub | |
| • Acaciella angustissima | ||
| var. filicioides (Cav.) L.Rico | shrub | |
| • Aeschynomene compacta | shrub | |
| Rose | shrub | |
| * Mimosa brevispicata | “uña de gato” | shrub |
| Britton | tree | |
| * Mimosa luisana Brandegee | ||
| *• Mimosa polyantha Benth. | “mantecoso” | tree |
| • Parkinsonia praecox (Ruiz & Pav. ex. Hook.) | “mezquite” | tree |
| Hawkins | ||
| • Prosopis laevigata (Humb., & Bonpl. ex Willd.) | “tecuahue” | shrub |
| M.C.Johnst. | ||
| • Senna wislizeni | ||
| var. pringlei (Rose) | ||
| H.S.Irwin & Barneby | ||
| Zapoteca formosa (Kunth) | ||
| H.M.Hern. | ||
| Heliotropiaceae | ||
| Heliotropium angiospermum | suffrutex | |
| Murray | ||
| Malpighiaceae | ||
| Calcicola parvifolia | shrub | |
| (A.Juss.) W.R.Anderson & C.Davis | ||
| Malvaceae | ||
| • Ceiba aesculifolia (Kunth) | “pochote” | tree |
| Britten & Baker f. | shrub | |
| Melochia tomentosa L. | ||
| Onagraceae | ||
| Ludwigia octovalvis (Jacq.) | herb | |
| P.H.Raven | ||
| Polygonaceae | ||
| Podopterus mexicanus | tree | |
| Bonpl. | ||
| Rhamnaceae | ||
| *• Ziziphus pedunculata | “cholulo” | shrub |
| (Brandegee) Standl. | ||
| Rubiaceae | ||
| Hintonia latiflora (DC.) | tree | |
| Bullock | “zapotillo” | shrub |
| • Randia nelsonii Greenm. | ||
| Sapindaceae | ||
| Cardiospermum | liana | |
| microcarpum Kunth | ||
| Solanaceae | ||
| * Solanum tridynamum | shrub | |
| Dunal | ||
| Talinaceae | ||
| Talinum paniculatum (Jacq.) | herb | |
| Gaertn. | ||
| Verbenaceae | ||
| Lantana camara L. | shrub | |
| • Lippia graveolens Kunth | “orégano” | shrub |

Figure 2 Most representative plants of tropical deciduous forest of San José Tilapa, Puebla. (a) Ceiba aesculifolia. (b) Bursera aptera. (c) Bursera morelensis. (d) Bursera submoniliformis. (e) Parthenium fruticosum. (f) Euphorbia schlechtendalii. (g) Fouquieria formosa. (h) Lippia graveolens. (i) Mimosa luisana. (j) Plumeria rubra. (k) Prosopis laevigata. (l) Ziziphus pedunculata.
Our survey showed that the abundance of life forms included 1,740 individuals (trees, shrubs, arborescent and rosette plants) recorded in the 220 quadrants of the 55 points marked in the 5 transects (Table 2).
Table 2 Richness (N), total counts in bold type and abundances of trees and shrubs, in parentheses abundances of these biological forms near to the center point in the 5 transects (in bold types).
| Transects | T1 | T2 | T3 | T4 | T5 | Total |
| N trees | 8 | 7 | 7 | 9 | 7 | N=13 |
| N shrubs | 4 | 5 | 4 | 8 | 5 | N = 9 |
| N arborescents | 0 | 1 | 1 | 1 | 1 | N = 4 |
| N rosettes | 1 | 0 | 1 | 1 | 1 | N = 4 |
| Total richness of trees and shrubs | 12 | 12 | 11 | 17 | 12 | N=22 |
| Abundance of trees | 58 | 73 | 49 | 75 | 125 | 380 |
| (37) | (38) | (34) | (39) | (42) | (190) | |
| Abundance of shrubs | 172 | 329 | 139 | 243 | 339 | 1,222 |
| (40) | (43) | (44) | (42) | (41) | (210) | |
| Abundance of arborescents | 0 | 13 | 1 | 20 | 33 | 67 |
| Abundance of rosettes | 21 | 0 | 11 | 2 | 37 | 71 |
| Total abundance of trees and shrubs | 230 | 402 | 188 | 318 | 464 | 1,602 |
| (77) | (81) | (78) | (81) | (83) | (400) | |
| Total abundance of all biological forms | 251 | 415 | 200 | 340 | 534 | 1,740 |
Exclusively considering the trees (190 individuals) and the shrubs (210 individuals) in all transects (T1-T5), a total of 22 species was recorded. In addition, transects T4 on the W slope showed the highest richness values (Table 2). Although the total abundance values showed a slight variation, transect T5 on the NW slope exhibited the highest total abundance, followed by transects T2 and T4 (Table 2).
The floristic composition of the TDF on El Picante hill comprised the tree species Bursera aptera, B. morelensis, Ceiba aesculifolia, Fouquieria formosa and Senna wislizeni var. pringlei. With the exception of B. morelensis, which was recorded in T1-T3 and T5, all of the other 4 species occurred in all 5 studied transects (T1-T5). The shrub stratum was made up of Mimosa polyantha, Lippia graveolens, Randia nelsonii and Parthenium fruticosum (T1, T3, T4 and T5). The first 2 species listed occurred in all 5 transects. It is also important to note that there were species unique to 4 of the 5 transects, as follows: Cyrtocarpa procera (T1), Acacia cochliacantha (T2), Ziziphus pedunculata (T4) and Bursera submoniliformis (T5). Similarly, the shrub stratum included Croton morifolius (T2), Cordia curassavica and Justicia candicans (T4), as taxa that exclusively occurred in these transects.
Estimating the vertical and horizontal structure
The analysis of the vegetation structure included 13 families, 20 genera and 22 species, all of them are marked with a symbol (•) in Table 1. The vertical data showed that species with the highest number of individuals in the tree stratum were Ceiba aesculifolia, followed by Bursera aptera and Fouquieria formosa; with respect to height, the medium group was the best represented (Fig. 3a-e). The most abundant taxa in the shrub stratum were Lippia graveolens, followed by Parthenium fruticosum and Mimosa polyantha. Most shrubs exhibited medium height (Fig. 3a-e).
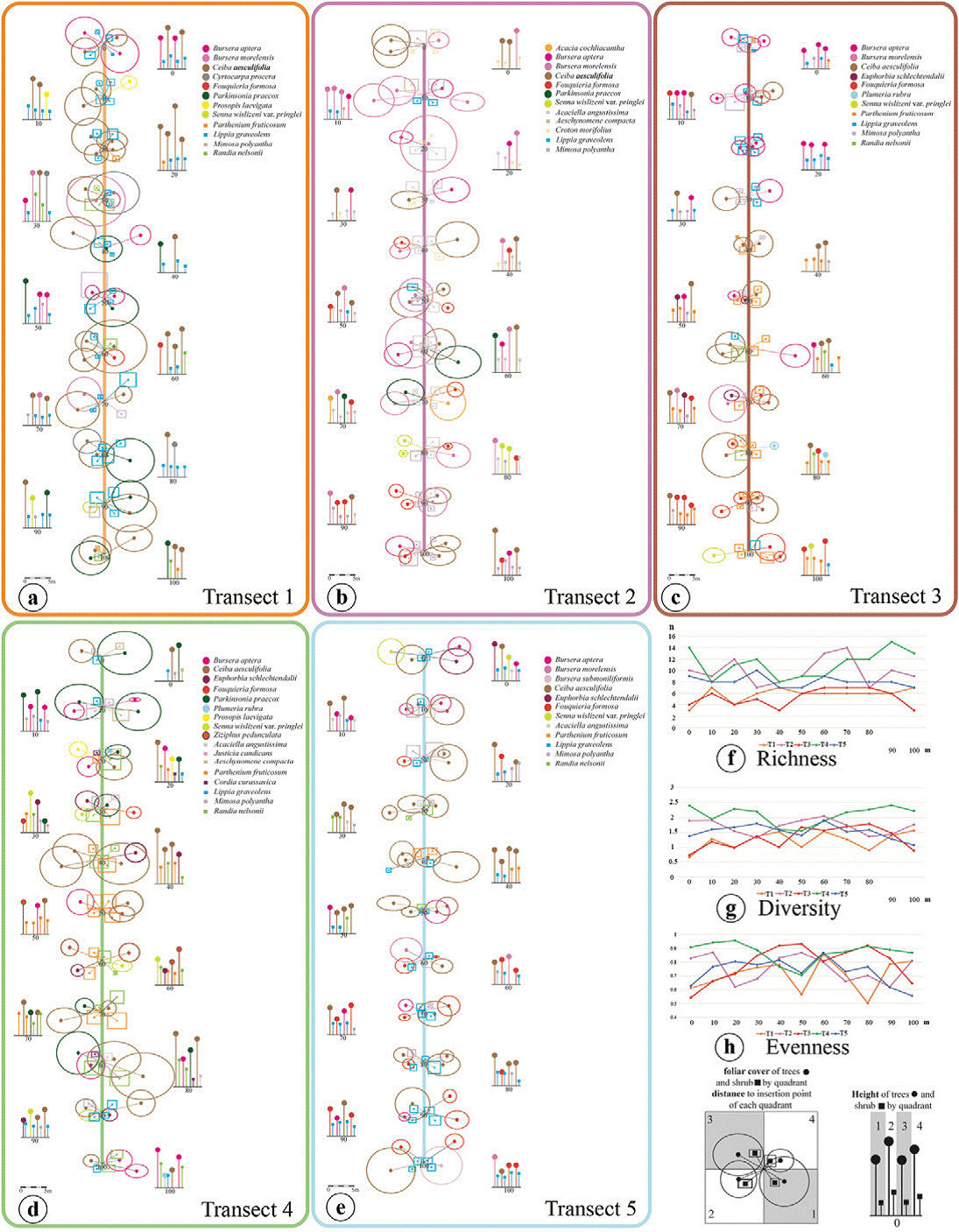
Figure 3 Schematic profiles of the vegetation cover of the trees and shrubs found in the 5 transects (a-e). Richness (f), diversity (g), and evenness (h) indices recorded in each transect.
About the horizontal structure, the total tree density (DenTha) of transects T1 and T3 recorded the highest values, while the largest basal area was found in transect T1. The species that showed the highest density were Bursera aptera, B. morelensis, Ceiba aesculifolia, Fouquieria formosa and Parkinsonia praecox. These species also showed the most significant IVI values. C. aesculifolia and B. aptera were recorded in 4 transects, F. formosa in 3, P. praecox in 2 and B. morelensis was only recorded in 1 transect (Table 3). As for the total density of the shrubs (DenTha), transects T3 and T5 (Fig. 3c, e) recorded the highest values, while the highest cover value was found in transect T4 (Fig. 3d). The shrubs with the highest densities were Lippia graveolens, Acaciella angustisima, Mimosa polyantha, Randia nelsonii, Croton morifolius and Parthenium fruticosum. These same taxa showed the higher IVI values (Table 4).
Table 3 Structural parameters of the species present in the tree stratum on the 5 transects in the TDF on El Picante hill, San José Tilapa, Mexico. Total density of individuals per hectare (DenTha), mean basal area (Ab), absolute density (DenA), relative density (DenR), absolute dominance (DomA), relative dominance (DomR), absolute frequency (FA), relative frequency (FR) and the importance value index (IVI). The 3 species with the highest IVI are indicated in bold type.
| N° | Species | DenTha | Ab | DenA | DenR | DomA | DomR | FA | FR | IVI (%) |
| 1 | Ceiba aesculifolia | 423.11 | 91.47 | 0.51 | 51.35 | 0.48 | 48.84 | 0.38 | 38.46 | 46.21 |
| Parkinsonia praecox | 111.34 | 27.87 | 0.14 | 13.51 | 0.14 | 14.88 | 0.19 | 19.23 | 15.87 | |
| Bursera aptera | 133.61 | 23.66 | 0.16 | 16.21 | 0.12 | 12.63 | 0.15 | 15.38 | 14.74 | |
| Bursera morelensis | 44.54 | 24.20 | 0.05 | 5.40 | 0.12 | 12.91 | 0.07 | 7.69 | 8.67 | |
| Cyrtocarpa procera | 44.54 | 15.13 | 0.05 | 5.40 | 0.08 | 8.08 | 0.07 | 7.69 | 7.05 | |
| Prosopis laevigata | 22.27 | 2.66 | 0.03 | 2.70 | 0.014 | 1.42 | 0.03 | 3.84 | 2.65 | |
| Fouquieria formosa | 22.27 | 1.57 | 0.03 | 2.70 | 0.008 | 0.84 | 0.03 | 3.84 | 2.46 | |
| Senna wislizeni var. pringlei | 22.27 | 0.69 | 0.03 | 2.70 | 0.003 | 0.37 | 0.03 | 3.84 | 2.30 | |
| Total | 823.94 | 187.28 | 1 | 100 | 1 | 100 | 1 | 100 | 100 | |
| 2 | Bursera morelensis | 150.04 | 50.08 | 0.28 | 28.94 | 0.52 | 52.5 | 0.29 | 29.03 | 36.82 |
| Ceiba aesculifolia | 136.4 | 22.62 | 0.26 | 26.31 | 0.23 | 23.71 | 0.22 | 22.58 | 24.2 | |
| Fouquieria formosa | 95.48 | 1.95 | 0.18 | 18.42 | 0.02 | 2.048 | 0.19 | 19.35 | 13.27 | |
| Bursera aptera | 68.2 | 9.86 | 0.13 | 13.15 | 0.103 | 10.33 | 0.16 | 16.12 | 13.2 | |
| Parkinsonia praecox | 27.28 | 9.82 | 0.05 | 5.26 | 0.102 | 10.3 | 0.06 | 6.45 | 7.33 | |
| Senna wislizeni var. pringlei | 27.28 | 0.49 | 0.05 | 5.26 | 0.005 | 0.51 | 0.03 | 3.22 | 3.001 | |
| Acacia cochliacantha | 13.64 | 0.56 | 0.02 | 2.63 | 0.0059 | 0.6 | 0.03 | 3.22 | 2.15 | |
| Total | 518.32 | 95.40 | 1 | 100 | 1 | 100 | 1 | 100 | 100 | |
| 3 | Ceiba aesculifolia | 247.11 | 31.32 | 0.29 | 29.41 | 0.36 | 35.86 | 0.33 | 33.33 | 32.87 |
| Bursera aptera | 271.82 | 30.66 | 0.32 | 32.35 | 0.35 | 35.10 | 0.25 | 25 | 30.82 | |
| Fouquieria formosa | 197.68 | 10.71 | 0.23 | 23.53 | 0.12 | 12.26 | 0.21 | 20.83 | 18.87 | |
| Bursera morelensis | 24.71 | 10.34 | 0.02 | 2.94 | 0.12 | 11.84 | 0.04 | 4.17 | 6.32 | |
| Euphorbia schlechtendalii | 49.42 | 2.76 | 0.05 | 5.88 | 0.03 | 3.16 | 0.08 | 8.33 | 5.79 | |
| Plumeria rubra | 24.71 | 0.78 | 0.02 | 2.94 | 0.01 | 0.89 | 0.04 | 4.17 | 2.67 | |
| Senna wislizeni var. pringlei | 24.71 | 0.78 | 0.02 | 2.94 | 0.01 | 0.89 | 0.04 | 4.17 | 2.67 | |
| Total | 840.16 | 87.36 | 1 | 100 | 1 | 100 | 1 | 100 | 100 | |
| 4 | Ceiba aesculifolia | 153.69 | 28.36 | 0.33 | 33.33 | 0.47 | 46.63 | 0.27 | 26.67 | 35.54 |
| Parkinsonia praecox | 82.75 | 16.00 | 0.17 | 17.95 | 0.26 | 26.30 | 0.20 | 20.00 | 21.42 | |
| Bursera aptera | 82.75 | 10.19 | 0.17 | 17.95 | 0.17 | 16.75 | 0.17 | 16.67 | 17.12 | |
| Euphorbia schlechtendalii | 35.47 | 0.95 | 0.07 | 7.69 | 0.02 | 1.57 | 0.10 | 10.00 | 6.42 | |
| Senna wislizeni var. pringlei | 35.47 | 0.79 | 0.07 | 7.69 | 0.01 | 1.30 | 0.10 | 10.00 | 6.33 | |
| Ziziphus pedunculata | 35.47 | 0.86 | 0.07 | 7.69 | 0.01 | 1.41 | 0.07 | 6.67 | 5.26 | |
| Fouquieria formosa | 11.82 | 3.34 | 0.02 | 2.56 | 0.05 | 5.50 | 0.03 | 3.33 | 3.80 | |
| Prosopis laevigata | 11.82 | 0.27 | 0.02 | 2.56 | 0.00 | 0.44 | 0.03 | 3.33 | 2.11 | |
| Plumeria rubra | 11.82 | 0.06 | 0.02 | 2.56 | 0.00 | 0.10 | 0.03 | 3.33 | 2.00 | |
| Total | 461.06 | 60.81 | 1 | 100 | 1 | 100 | 1 | 100 | 100 | |
| 5 | Ceiba aesculifolia | 290.13 | 33.68 | 0.52 | 52.38 | 0.62 | 62.36 | 0.42 | 42.31 | 52.35 |
| Fouquieria formosa | 131.88 | 5.45 | 0.23 | 23.81 | 0.10 | 10.10 | 0.23 | 23.08 | 19.00 | |
| Bursera aptera | 79.13 | 7.88 | 0.14 | 14.29 | 0.15 | 14.59 | 0.19 | 19.23 | 16.04 | |
| Bursera submoniliformis | 13.19 | 3.73 | 0.02 | 2.38 | 0.07 | 6.91 | 0.04 | 3.85 | 4.38 | |
| Bursera morelensis | 13.19 | 1.66 | 0.02 | 2.38 | 0.03 | 3.07 | 0.04 | 3.85 | 3.10 | |
| Euphorbia schlechtendalii | 13.19 | 1.27 | 0.02 | 2.38 | 0.02 | 2.35 | 0.04 | 3.85 | 2.86 | |
| Senna wislizeni var. pringlei | 13.19 | 0.34 | 0.02 | 2.38 | 0.01 | 0.62 | 0.04 | 3.85 | 2.28 | |
| Total | 553.88 | 54.00 | 1 | 100 | 1 | 100 | 1 | 100 | 100 |
Table 4 Structural parameters of the species present in the shrub stratum, on the 5 transects in the TDF on El Picante hill, San José Tilapa, Mexico. Density of individuals per hectare (DenTha), mean foliar cover (Cob), absolute density (DenA), relative density (DenR), dominance (DomA), relative dominance (DomR), absolute frequency (FA), relative frequency (FR) and the importance value index (IVI). The 3 species with the highest IVI are indicated in bold type.
| N° | Species | DenTha | Cob | DenA | DenR | DomA | DomR | FA | FR | IVI (100%) |
| 1 | Lippia graveolens | 1,176.53 | 1,837.95 | 0.67 | 67.5 | 0.44 | 44.10 | 0.53 | 52.63 | 54.74 |
| Mimosa polyantha | 305.03 | 1,589.39 | 0.17 | 17.5 | 0.38 | 38.13 | 0.21 | 21.05 | 25.56 | |
| Randia nelsonii | 174.30 | 675.37 | 0.1 | 10 | 0.16 | 16.20 | 0.16 | 15.79 | 14 | |
| Parthenium fruticosum | 87.15 | 65.40 | 0.05 | 5 | 0.02 | 1.57 | 0.11 | 10.53 | 5.70 | |
| Total | 1,743 | 4,168.14 | 1 | 100 | 1 | 100 | 1 | 100 | 100 | |
| 2 | Acaciella angustissima | 914.67 | 2,507.88 | 0.44 | 44.19 | 0.47 | 47.02 | 0.33 | 33.33 | 41.51 |
| Mimosa polyantha | 625.83 | 2,069.50 | 0.30 | 30.23 | 0.39 | 38.80 | 0.33 | 33.33 | 34.12 | |
| Croton morifolius | 336.99 | 290.69 | 0.16 | 16.28 | 0.05 | 5.45 | 0.19 | 19.05 | 13.59 | |
| Lippia graveolens | 144.42 | 230.26 | 0.06 | 6.98 | 0.04 | 4.32 | 0.10 | 9.52 | 6.94 | |
| Aeschynomene compacta | 48.14 | 234.80 | 0.02 | 2.33 | 0.04 | 4.40 | 0.05 | 4.76 | 3.83 | |
| Total | 2,070.05 | 5,333.13 | 1 | 100 | 1 | 100 | 1 | 100 | 100 | |
| 3 | Parthenium fruticosum | 1,293.46 | 2,223.29 | 0.47 | 47.73 | 0.43 | 42.64 | 0.36 | 36.36 | 42.24 |
| Lippia graveolens | 739.12 | 1,240.36 | 0.27 | 27.27 | 0.24 | 23.79 | 0.27 | 27.27 | 26.11 | |
| Mimosa polyantha | 554.34 | 1,158.68 | 0.20 | 20.45 | 0.22 | 22.22 | 0.27 | 27.27 | 23.32 | |
| Randia nelsonii | 123.19 | 591.38 | 0.04 | 4.55 | 0.11 | 11.34 | 0.09 | 9.09 | 8.33 | |
| Total | 2,710.11 | 5,213.72 | 1 | 100 | 1 | 100 | 1 | 100 | 100 | |
| 4 | Randia nelsonii | 583.48 | 2,924.99 | 0.28 | 28.57 | 0.389 | 38.91 | 0.26 | 25.93 | 31.13 |
| Parthenium fruticosum | 583.48 | 2,916.76 | 0.28 | 28.57 | 0.388 | 38.80 | 0.19 | 18.52 | 28.63 | |
| Lippia graveolens | 340.36 | 536.55 | 0.16 | 16.67 | 0.071 | 7.14 | 0.19 | 18.52 | 14.11 | |
| Mimosa polyantha | 243.12 | 587.88 | 0.11 | 11.90 | 0.078 | 7.82 | 0.15 | 14.81 | 11.51 | |
| Aeschynomene compacta | 97.25 | 325.37 | 0.04 | 4.76 | 0.043 | 4.33 | 0.07 | 7.41 | 5.50 | |
| Cordia curassavica | 97.25 | 90.68 | 0.04 | 4.76 | 0.012 | 1.21 | 0.07 | 7.41 | 4.46 | |
| Acaciella angustissima | 48.62 | 89.29 | 0.02 | 2.38 | 0.012 | 1.19 | 0.04 | 3.70 | 2.42 | |
| Justicia candicans | 48.62 | 46.74 | 0.02 | 2.38 | 0.006 | 0.62 | 0.04 | 3.70 | 2.24 | |
| Total | 2,042.18 | 7,518.25 | 1 | 100 | 1 | 100 | 1 | 100 | 100 | |
| 5 | Lippia graveolens | 2,130.68 | 3,177.43 | 0.61 | 61.90 | 0.52 | 52.28 | 0.45 | 45.45 | 53.21 |
| Mimosa polyantha | 573.64 | 1,763.05 | 0.16 | 16.67 | 0.29 | 29.01 | 0.27 | 27.27 | 24.32 | |
| Acaciella angustissima | 327.80 | 419.76 | 0.09 | 9.52 | 0.07 | 6.91 | 0.14 | 13.64 | 10.02 | |
| Randia nelsonii | 245.85 | 418.62 | 0.07 | 7.14 | 0.07 | 6.89 | 0.09 | 9.09 | 7.71 | |
| Parthenium fruticosum | 163.90 | 298.44 | 0.04 | 4.76 | 0.05 | 4.91 | 0.05 | 4.55 | 4.74 | |
| Total | 3,441.87 | 6,077.29 | 1 | 100 | 1 | 100 | 1 | 100 | 100 |
In the TDF on El Picante hill, most of the trees exhibited a height in the range of 4 to 6 metres, placing them in the medium group. Both the short and the tall trees followed in importance. The shrub stratum showed dense canopy, more than 50% of the individuals were of medium height (Fig. 3b, c).
The analysis of the height and canopy cover data for the species present in the tree and shrub strata in the 5 transects (Fig. 3a-e) showed that Ceiba aesculifolia was the tallest tree (2.60-12.0 m) and it also exhibited the largest canopy cover (small canopy cover or c = 1.6-11.0; large canopy cover or C = 2.4-12.8 m). Randia nelsonii, present in transects T1, T3 and T4 (Fig. 3a, c, d), was the tallest shrub (0.9-7.0 m) and it also had the greatest canopy cover (c = 0.64-4.2; C = 0.7-4.3 m). Another species that is important to mention for transects T1, T3 and T5 (Fig. 3a, c, e) is Mimosa polyantha which exhibited high canopy cover values (c = 0.67-4.84; C = 0.7-5.05 m). Acaciella angustissima was significant in T2 (Fig. 3b) as it showed the greatest foliar cover (c = 0.65-6.7; C = 0.74-6.9 m).
Moreover, the results of this study allowed us to draw some interesting deductions; transects T1 (at the top of the hill) and T4 (to the western) exhibited the largest canopy cover, within the tree stratum highlighted B. morelensis, C. aesculifolia, C. procera and P. praecox, all of them were also the tallest trees. Meanwhile, in the shrub stratum M. polyantha, R. nelsonni and Z. pedunculata recorded the most important and the tallest shrubs. All transect showed very similar richness values of trees and shrubs with exception of T4 which recorded the highest richness value (Table 2, Fig. 3). On the other hand, T5 stood out by the presence of the greatest abundance of trees and shrubs (Table 2).
Ecological parameters
The highest diversity values were observed in T4 (Fig. 3d). The former transect recorded the highest number of species and it is where the abundance of organisms was most equitably distributed among the species, while the latter also showed a high richness value but not in terms of evenness (Fig. 3f, h).
The Jaccard index showed that transects T3 and T5 had the highest percentages of similarity (Table 5). The species shared between these transects were Bursera aptera, B. morelensis, Ceiba aesculifolia, Euphorbia schlechtendalii, Fouquieria formosa, Senna wislizeni var. pringlei, L. graveolens, Mimosa polyantha, Parthenium fruticosum, and Randia nelsonii. On the other hand, Plumeria rubra was exclusive to the transect T3, and both Acaciella angustissima and Bursera submoniliformis were unique to the transect T5.
Table 5 Jaccard similarity indices among the transects (T1-T5): in parentheses, the total number of species compared and the shared species are indicated, the highest value is in bold type.
| Transects | T1 | T2 | T3 | T4 | T5 |
| T1 | - | ||||
| T2 | 50.0% | (16/8) | - | ||
| T3 | 64.3% | (14/9) | 43.8% (16/7) | - | |
| T4 | 52.6% | (19/10) | 45.0% (20/9) | 55.6% (18/10) | - |
| T5 | 60% (15/9) | 50.0% (16/8) | 76.9% (13/10) | 52.6% (19/10) | - |
Discussion
This study focused exclusively on tree and shrub strata in the TDF on El Picante hill in San José Tilapa (Coxcatlán), Puebla, within the Tehuacán Valley. Although collection was performed among the transects in order to supplement the floristic inventory, the total richness observed was low compared with those studies performed within plots (Table 6). However, it should be mentioned that in another research conducted in the same municipality of Coxcatlán, in the town of Calipan, Trejo (2005) whom did not explain the methods of sampling, also reported low species richness (29 spp.), which she justified by the study area proximity to other types of vegetation with an affinity towards xeric environments, such as xerophilous scrub (Table 6).
Table 6 Comparative table including richness (N) and the Shannon-Wiener (H’) and Pielou (J’) indices recorded in studies of structure and floristic composition of the Mexican tropical deciduous forest. Values recorded in this study are in bold type.
| N | H’ | H’ | J’ | J’ | |
| range | range | average | range | average | |
| Gallardo-Cruz et al. (2005) (plots) | 21- 39 | 2.12 - 3.16 | - | 0.67 - 0.89 | - |
| Rocha-Loredo et al. (2010) (plots) | 13-14 | - | 2.1 ± 0.07 | - | 0.83 ± 0.02 |
| - | 2.3 ± 0.017 | - | 0.86 ± 0.05 | ||
| Dzib-Castillo et al. (2014) (plots) | 6-15 | 0.94 - 2.24 | 1.91 | 0.43 - 0.84 | 0.63 |
| Méndez-Toribio et al. (2014) (plots) | 7-56 | 0.41 - 2.69 | 1.98 | - | - |
| Trejo (2005) | 29 | - | 2.96 | - | 0.88 |
| This study (transects) | 12- 22 | 0.67 - 2.42 | 2.07 | 0.71 - 0.87 | 0.74 |
Although the tropical deciduous forest we studied in San José Tilapa did not exhibit a high level of species richness, our physiognomic analysis revealed the presence of taxa that are characteristic of this plant community, such as neotropical plant families and some genera such as Apocynaceae (Plumeria), Cordiaceae (Cordia), Burseraceae (Bursera), Cactaceae (Myrtillocactus, Pachycereus, and Opuntia), Euphorbiaceae (Euphorbia and Croton), Fabaceae (Acacia, Mimosa, Parkinsonia, Senna), and Malvaceae (Ceiba), which coincides with previous research conducted in other tropical deciduous forests of Mexico (Bravo-Bolaños et al., 2016; Gallardo-Cruz et al., 2005; León-de la Luz et al., 2012; Lott & Atkinson, 2010; Méndez-Toribio et al., 2014; Pineda-García et al., 2007; Rzedowski, 1978; Sánchez-Mejía et al., 2007; Sánchez-Velásquez et al., 2002; Sousa, 2010; Trejo, 2010). These families have also been important in the secondary communities of the TDF (Beltrán-Rodríguez et al., 2018).
The Fabaceae family has not only been classified as one of the most diverse families, but it also was abundantly represented in these plant communities. A high number of individuals were recorded in this study (n = 90), which is in agreement with reports published by Gallardo-Cruz et al. (2005) and Méndez-Toribio et al. (2014). Although these authors conducted their studies using different methods (Table 6), they recorded a high number of individuals from the Fabaceae family. According to Rzedowski and Calderón-de Rzedowski (2013), species of the genus Acacia and Mimosa that occur in San José Tilapa are exclusive or preferent of the TDF. Other taxa that are distinctive to the TDF studied were Cordia (Cordiaceae), Euphorbia (Euphorbiaceae), and Randia (Rubiaceae), which are genera reported by Rzedowski and Calderón-de Rzedowski (2013).
It is well-documented that the Cactaceae family, which occurs in Mexico’s tropical deciduous forests, places its characteristic seal on the dry season with its various biological forms (Rzedowski, 1978; Trejo, 1999, 2005), and the physiognomy of the TDF on El Picante hill is no exception: there were arborescent life forms (Opuntia) quantified in 4 transects, along with branched columnar cacti (Myrtillocactus geometrizans and Pachycereus weberi) and globular cacti (Coryphantha pallida) present in the vicinity of the sampling sites. This coincides with the research of Bravo-Bolaños et al. (2016) in the TDF of the coastal region of Banderas Bay in Nayarit, who reported the presence of cacti of the genera Pachycereus and Opuntia. Another life form that enriched the physiognomy we observed were the rosettes, represented by the genus Agave (Asparagaceae) and absent only in the T2 transect.
The physiognomy of the TDF was characterised by trees that varied in height from 4 to 15 m. However, their mean height ranges from 8 to 12 m, and some exceptional isolated life forms that reach heights of ca. 15 m are present in this community (Challenger & Soberón, 2008; Miranda & Hernández, 1963; Pennington & Sarukhán, 2005; Rzedowski, 1978; Trejo, 1998). With regard to height and canopy cover, among the trees and shrubs of the TDF under study, Ceiba aesculifolia, Mimosa polyantha and Randia nelsonii, respectively, were the most notable.
Analysis of the horizontal structure of the tree stratum in the tropical deciduous forest revealed 5 species that showed relatively high importance indices (IVIs): Ceiba aesculifolia, Bursera morelensis, B. aptera, Parkinsonia praecox, and Fouquieria formosa. The results of this study coincide with 7 species reported by Méndez-Toribio et al. (2014), whose research was conducted in Tziritzícuaro, Balsas Depression, Michoacán. However, there are differences in the IVI, since Méndez-Toribio et al. (2014) reported low values for C. aesculifolia, while Acacia cochliacantha, Cyrtocarpa procera, Euphorbia schlechtendalii, Plumeria rubra (4.33%), and Randia nelsonii recorded higher IVI values than those obtained in the TDF of San José Tilapa.
Bravo-Bolaños et al. (2016) reported various tree and shrub genera with high IVI values such as Acacia, Bursera, Enterolobium, Eysenhardtia, Ficus and Plumeria from the tree stratum, while for the shrub stratum Bauhinia, Celtis, Mimosa, Opuntia, Randia and Ziziphus are mentioned; unfortunately, the values were not specified. Furthermore, Beltrán-Rodríguez et al. (2018) reported a lower value for Euphorbia schlechtendalii (4.8%) and a higher value for Acacia cochliacantha (4.7%) than those recorded in this study.
Upon comparison of the diversity and evenness indices reported in other studies of Mexican tropical deciduous forests (Dzib-Castillo et al., 2014; Gallardo-Cruz et al., 2005; Méndez-Toribio et al., 2014; Rocha-Loredo et al., 2010), with the values found in San José Tilapa, Puebla, in this study (Table 6), it is clear that the values reported are very similar, with the exception of the research performed in Oaxaca by Gallardo-Cruz et al. (2005), which found a higher diversity index is shown (H’ = 3.16).
Severe climate changes have been affected plant communities (Barnosky et al., 2011; Ceballos, García et al., 2010; Ceballos, Martínez et al., 2010; Ceballos et al., 2015). This alteration has caused such an impact in the distribution and phenology of terrestrial ecosystems (Chmura et al., 2019; Donnelly & O’Neill, 2013; Gordo & Sanz, 2010; Richardson et al., 2013; Workie & Debella, 2018). The tropical deciduous forest (TDF) is one of the most significant ecological components at the national level that has been severely altered. Moreover, TDF is well represented at Tehuacán Valley, today considered a megadiverse region with a high level of endemism (Dávila et al., 1993, 2002; Téllez-Valdés et al., 2008; Valiente-Banuet et al., 2000, 2009). Overall, our study documents a low floristic richness of trees and shrubs, probably of an early secondary TDF at the Tehuacán Valley (Mesa-Sierra et al., 2020).
Among the most relevant signs of this study, that must be applied not only in this locality, but also throughout all Tehuacán-Cuicatlán Valley, are the dramatic changes in land use and the effects of climate change on the plant communities. On one hand, this research at El Picante hill provides additional knowledge about TDF distribution, its composition and structure in this valley, and on the other hand, it shows us the need to protect this ecosystem wherever it is less diverse and perhaps more exposed due to human activities that take place in San José Tilapa. We believe that this contribution will enable the community to issue recommendations on the protection of this vulnerable type of vegetation and in the future, it will be important to evaluate the most vulnerable native plant species to human activities and climate change that unfortunately could trigger on habitat fragmentations.

