











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
La ecología demográfica estudia la variación del tamaño y densidad de una población en tiempo y espacio. A nivel temporal, se analiza la dinámica poblacional de una especie al concentrar estadísticamente los parámetros demográficos como supervivencia, fecundidad y crecimiento poblacional, que expresan en promedio el comportamiento de una población en un intervalo de tiempo determinado (Larson-Guerra, 1992; Caswell, 2001). El análisis de estos parámetros en modelos matriciales permite describir la demografía y, con ello, determinar la tasa de crecimiento poblacional y analizar qué parámetros son más sensibles a cambios (Harper, 1977; Caswell, 2001). Por ello, los modelos matriciales son una herramienta que proporciona resultados importantes y útiles para el diseño de estrategias de conservación (Alfonso-Corrado, Clark-Tapia y Mendoza, 2007; Ezard et al., 2010; Primack, 2012) y manejo (Alfonso-Corrado et al., 2007). En especies arbóreas amenazadas, endémicas, o relicto, estos modelos permiten determinar el estado actual de sus poblaciones y predecir su comportamiento a largo plazo bajos diferentes escenarios antropogénicos (e.g., cambio de uso de suelo, manejo forestal, etc.) o naturales (i.e., cambio climático) (Ezard et al., 2010; Primack, 2012; You, Liu y Fujiwara, 2013). Por ejemplo, estudios demográficos realizados en especies relicto (Hampe y Arroyo, 2002; Picó y Ribal, 2002; García, 2003) han mostrado que las poblaciones de Frangula alnus sub. baetica Mill, Borderea chouardii (Gaussen), Ramonda myconi (L). R.chb. son dominadas por individuos adultos y senescentes y, a pesar que presentan serios problemas en el reclutamiento debido a la alta mortalidad de plántulas, sus poblaciones son estables demográficamente, con valores de lambda cercanos o iguales a la unidad. Sin embargo, García (2003) encontró, para Borderea chouardii (Gaussen), alta susceptibilidad a disturbios, con baja capacidad de recuperación, aspecto similar encontrado en otras especies amenazadas (Clark-Tapia, Alfonso-Corrado, Mandujano y Molina-Freaner, 2006; You et al., 2013).
Oreomunnea mexicana (Standl.) J. F. Leroy es una especie de árbol relicto que surgió en el Mioceno tardío y Plioceno temprano hace aproximadamente 23 millones de años (Palacio-Chávez y Rzendowski, 1993); actualmente está catalogada como amenazada (González-Espinoza et al., 2011; Alfonso-Corrado et al., 2017), pertenece a la familia Juglandaceae y es la única especie representante del género Oreomunnea en México (Russell y Cohn, 2012). Su distribución es fragmentada, a lo largo de los bosques de niebla de México y Centroamérica, y forma poblaciones monodominadas por grandes individuos de O. mexicana, en los lugares en que ocurre (Navare, 1983; Corrales, Arnold, Ferrer, Turner y Dalling et al., 2016; Alfonso-Corrado et al., 2017). Su distribución en México se restringe a los estados de Veracruz, y Oaxaca (González-Espinoza et al., 2011), siendo en este último estado, donde se encuentran las poblaciones más extensas de la especie (Naranjo-Luna, 2014), en el bosque de niebla más grande y conservado de México (Anta-Fonseca et al., 2010). En esta región, O. mexicana es considerada una especie bandera de importancia cultural y biológica para las comunidades chinantecas de la Sierra Juárez, por lo que el conocimiento de sus aspectos biológicos es fundamental no solo para su conservación, sino también para la del hábitat del bosque nuboso (Naranjo-Luna, 2014; Alfonso-Corrado et al., 2017).
Estudios genéticos recientes realizados en la Sierra Juárez, Oaxaca, sugieren que las poblaciones de la especie no se encuentran depauperadas (Naranjo-Luna, 2014; Pascual-Mendoza, 2014), sin embargo, un estudio ecológico realizado por Alfonso-Corrado et al. (2017) indica que la especie es altamente susceptible a procesos de cambio climático y modificación del nicho ecológico. Una forma precisa de identificar y analizar la estabilidad y riesgo de permanencia de O. mexicana es estudiar su dinámica poblacional a través del uso de modelos matriciales de proyección poblacional (Caswell, 2001) y con base en estos resultados sugerir acciones de conservación in situ para la especie.
Objetivos
Generar conocimiento biológico de O. mexicana, una especie eje, relicto y amenazada del bosque de niebla de Santiago Comaltepec, por medio del análisis de la estructura y dinámica poblacional, con la finalidad de proponer estrategias de conservación in situ de la especie.
Materiales y métodos
Sitio de estudio
El estudio demográfico se realizó en una población de O. mexicana, conocida como San Bernardo (17°36’47.45" de latitud norte y 96°22’37.72" de longitud oeste) (Naranjo-Luna, 2014), localizada en el bosque de niebla ubicado en municipio de Santiago Comaltepec, en la Sierra Juárez de Oaxaca, al sur de México (Fig. 1). El sitio tiene una elevación de 1750 m snm, orientación noroeste y pendiente de 30°. La temperatura y precipitación media anual son 15 ºC y 700 mm respectivamente (Anta-Fonseca et al., 2010).

Especie de estudio
Oreomunnea mexicana (Standl.) J.F. Leroy (nombre común: caudillo; nombre en Chinanteco: maá hue loó; que significa árbol que se descascara) es un árbol relicto del mioceno tardío y plioceno temprano de aproximadamente 23 millones de años del orden Fagales (Rzedowski y Palacio-Chávez, 1977; Palacio-Chávez y Rzendowski, 1993), se distribuye de forma discontinua en México en los estados de Veracruz y Oaxaca (González-Espinoza et al., 2011). En Oaxaca es la especie dominante en una variante de bosque de niebla con árboles de 30 m a 60 m de altura, entre 1000 m y 2000 m snm en lugares húmedos en la Sierra Juárez y Mazateca (Rzedowski y Palacio-Chávez, 1977; Torres-Colín, 2004).
La especie presenta hojas perennes pinnadas desde cuatro hasta ocho foliolos y dispuestas en pares opuestos con una coloración blanca en el envés. El fruto es una nuez globosa encerrada por la base de la bráctea abaxial y el lóbulo bracteolar adaxial, de 0.8 cm - 1.1 cm de longitud y 0.6 cm - 0.8 cm de ancho, la superficie con escamas peltadas amarillas y pardas. La corteza exfoliante que se desprende en pedazos grandes y el fruto con una larga bráctea trilobada son características importantes para distinguir a la especie (Russell y Cohn, 2012).
Selección de sitio y trabajo de campo
En agosto de 2011 se seleccionaron cuatro sitios (Cerro Hormiga, El Panteón, La Cañada y San Bernardo) con presencia de O. mexicana para analizar su estructura poblacional (Fig.1). Dentro de los sitios se estableció una parcela de 400 m2 (20 m × 20 m), en la que se marcaron con etiqueta a todos los individuos de O. mexicana. La dimensión de la parcela fue con la finalidad de obtener un mínimo de 10 individuos adultos, basada en la experiencia en campo y en las recomendaciones de Samo-Lumbreras, Garmendia-Salvador y Delgado, (2008). A los individuos pequeños que no presentaban un tallo lignificado se les registró el diámetro basal, mientras que a los adultos con tallo lignificado se les registró el diámetro a la altura del pecho (DAP). Solamente se analizó la dinámica poblacional de San Bernando pues, debido a problemas de logística, no fue posible analizar todos los sitios. En San Bernardo todos los individuos marcados en agosto de 2011, fueron medidos nuevamente en septiembre de 2015, registrando crecimiento, supervivencia, mortalidad y nuevos reclutas.
El 9 marzo de 2016, se registraron en la zona de estudio fuertes vientos con velocidad superior a los 100 km/h (Clark-Tapia, Suárez-Mota, Nolasco y Velasco-Hipólito, 2016), que afectaron severamente a diversas poblaciones de O. mexicana. Por ello, en septiembre de 2016 se censó nuevamente la población de San Bernando. Además, se registró el daño en los individuos localizados dentro de las parcelas establecidas en 2011 de Cerro Hormiga, El Panteón y La Cañada, en estos sitios solamente se evaluaron cambios en la estructura poblacional. Los individuos registrados en los cuatro sitios fueron clasificados en cuatro categorías de tamaño, de acuerdo con su diámetro a la altura de pecho (DAP) o con su diámetro basal: 1) 0 cm - 5 cm, 2) 5.1 cm - 10 cm, 3) 10.1 cm - 40.0 cm y 4) > 40.1 cm.
Análisis Matricial
Se utilizó un modelo matricial tipo Lefvovich (Lefkovich, 1965; Caswell, 2001) el cual se describe en la siguiente ecuación:
El modelo matricial de Lefkovich está formado por un vector columna (n) que contiene el número de individuos en cada categoría de tamaño, al tiempo t y t+1, y una matriz A . La matriz A está conformada por diferentes elementos del ciclo de vida de una especie que definen las probabilidades de: 1) supervivencia, referida a la probabilidad que un individuo sobreviva o permanezca en la misma categoría (se ubican en la diagonal principal de la matriz), 2) crecimiento, descrito como la probabilidad de que un individuo pase a la siguiente categoría de tamaño (se ubica inmediatamente debajo de la diagonal principal de la matriz), 3) retroceso, representado como la probabilidad que un individuo retroceda de un estadio a otro (en la matriz se ubica inmediatamente arriba de la diagonal principal) y 4) fecundidad, que constituye la contribución que hace un individuo de una categoría en el tiempo t a cada una de las diferentes categorías en el intervalo del tiempo t+1 (se ubica en el primer renglón de la matriz).
Para la especie en estudio, se construyeron dos matrices tipo Lefkovich de dimensiones de 4 × 4; la primera del período que correspondió al tiempo inicial t en 2011 y t+1 al 2015, y la segunda fue del tiempo t (2015) al tiempo t+1 ( 2016). El ánalisis matricial se realizó con el software Pooptools versión 3.2.5 (Hood, 2010), con el cual se obtuvo la tasa finita de crecimiento poblacional (λ) y la estructura estable de tamaño (w). Para estimar el intervalo de confianza de λ se usó el método analítico de acuerdo con Caswell (2001).
Análisis de elasticidad
Se realizó un análisis de elasticidad desarrollado por de Kroon, Plaisier, van Groenendael y Caswell (1986), para los períodos (2011-2015 y 2015-2016). Dicho análisis calcula el cambio infinitesimal proporcional en las entradas de la matriz de transición de Lefkovich arriba mencionadas, en otras palabras, permite comparar la importancia relativa de los diferentes parámetros demográficos (supervivencia, crecimiento, retroceso y fecundidad) o de las diferentes categorías de tamaño, sobre la tasa finita de crecimiento poblacional obtenida en la matriz del período 2011-2015 y 2015-2016.
Simulaciones de cosecha
Aunque O. mexicana no tiene actualmente uso forestal local; se exploró la capacidad de la población de mantener el equilibrio demográfico. Por lo anterior, se realizaron simulaciones de cosecha de 5%, 10%, 15%, 25%, 50%, 80% y 100% orientada a extraer los individuos adultos con un tallo bien desarrollado (categorías 3 y 4) y que presentaron fecundidad, para determinar el porcentaje de cosecha por la cual la tasa de crecimiento poblacional (λ) cae por debajo de la unidad, el análisis se realizó solamente en la matriz del período 2011 al 2015.
Resultados
La estructura poblacional mostró un comportamiento en forma de J invertida en los cuatro sitios analizados, siendo la primera categoría la que mostró significativamente una mayor abundancia de individuos (q = 7.87, p < 0.05), mientras que la categoría 4 tuvo la menor abundancia (Fig. 2). Este patrón, se mantuvo en 2016, a pesar de que en este año se registró una alta mortalidad de individuos en todas las categorías de tamaño. El decremento de individuos fue más severo en tres de las cuatro poblaciones (San Bernardo, El Panteón y La Cañada); en ellas murió más de 50% de los individuos de cada población, en contraste con Cerro Hormiga que presentó una mortalidad mínima, y poco impacto del evento meteorológico extremo descrito en métodos (Fig. 2).
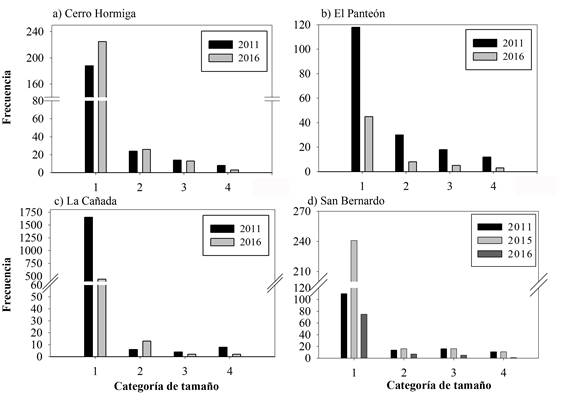
Figura 2 Número de individuos por categoría de tamaño de las poblaciones estudiadas en Santiago Comaltepec, durante 2011, 2015 y 2016.
El análisis demográfico realizado en el período de 2011 a 2015 a la población de San Bernardo, se encontró una tasa finita de crecimiento poblacional (λ) igual a 1.101, con un intervalo de confianza de λ ( 0.018. La simulación de cosecha de individuos adultos realizada a esta población registró una disminución inferior a la unidad en el valor de lambda (λ), cuando se extrajo en conjunto más de 25% de los individuos de las categorías 3 y 4 (Fig. 3). En contraste, cuando se cosecharon individuos de solamente una categoría (3 ó 4), la disminución de λ fue menos drástica, ya que se requirió de una extracción superior a 50% de los individuos (Fig. 3).
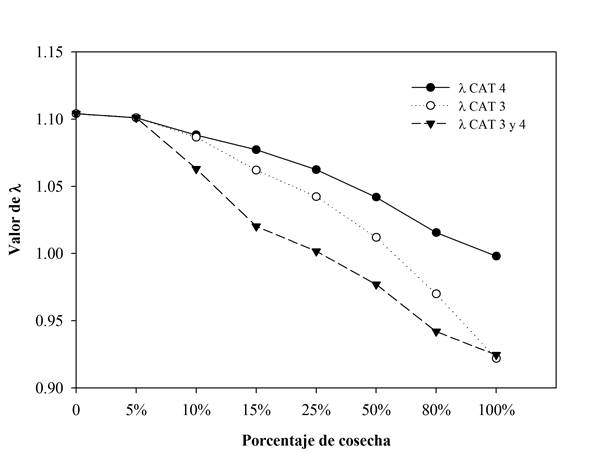
Figura 3 Cambios en la tasa de crecimiento poblacional (λ) en la población de San Bernardo después de una extracción de individuos (cosecha) en las categorías 3 y 4, y de una extracción en ambas categorías.
El resultado anterior indicó que la extracción de individuos en más de una categoría tuvo un efecto negativo en el crecimiento poblacional de O. mexicana. Esto se corroboró en el año 2016, donde la muerte por caída de árboles (> 60% de la población) afectó severamente diversas categorías de tamaño, ocasionando un decremento en la tasa finita de crecimiento de la población de San Bernardo (λ = 0.44±0.009). Un decremento similar en la tasa de crecimiento poblacional se infirió para las poblaciones de La Cañada y El Panteón donde murieron más de 70% y 65% de los individuos, respectivamente; mientras que la población de Cerro Hormiga permaneció estable o en crecimiento, debido a que se registró un leve incremento en el número de individuos (Fig. 3).
El análisis demográfico en San Bernardo mostró que la estructura poblacional observada y la predicha por el modelo en ambos períodos de estudio difieren significativamente (G 2= 12.59, p<0.001). En el período 2011-2015, para la categoría 1, la proporción de individuos observada fue menor que la esperada; mientras que en las categorías restantes la proporción observada fue mayor a la esperada (Fig. 4). En el período 2015-2016, la frecuencia esperada fue positiva solo en la categoría 3, donde se obtuvo una proporción máxima (Fig. 4).
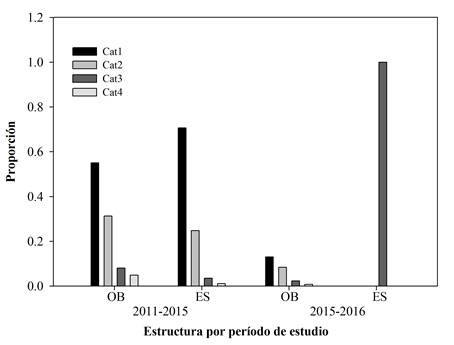
Figura 4 Estructura poblacional observada (OB) y esperada (ES) en las categorías de tamaño para la población de San Bernardo durante los períodos 2011-2015 y 2015-2016.
El análisis de elasticidad para el período 2011-2015 se muestra en la figura 5, este análisis reveló que la permanencia (P) fue el parámetro demográfico con mayor contribución relativa al valor de λ, seguida de la transición (T) y de la fecundidad (F). En contraste, para el período 2015-2016, la única contribución relativa al valor de λ fue dada por la permanencia de la categoría 3 (Fig. 5).

Discusión
El tamaño poblacional o la densidad poblacional en especies relictos o endémicas es un factor que se ha generalizado para determinar el riesgo de extinción y persistencia de una especie (Lande, 1993; Boyce, Haridas y Lee, 2006). Sin embargo, la presencia estable de poblaciones fragmentadas con densidades pequeñas en especies de este tipo, contradicen esta hipótesis y su decremento poblacional es difícil de detectar en una escala de estudio a corto plazo (García, 2008). Al factor de tamaño poblacional, deben sumarse los procesos de variabilidad ambiental y actividades antropogénicas que afectan los parámetros demográficos de una especie (Boyce et al., 2006); esto es particularmente importante en especies amenazadas, endémicas o relicto, cuyas poblaciones son más sensibles a procesos de cambio ambiental o disturbios (Naranjo-Luna, 2014), como lo es O. mexicana.
Oreomunnea mexicana es una especie relicto amenazada (Rzedowski y Palacio-Chávez, 1977; Palacio-Chávez y Rzendowski, 1993; González-Espinoza et al. 2011) con alta sensibilidad a procesos de cambio ambiental y una alta especificidad al hábitat donde se distribuye (Naranjo-Luna, 2014; Alfonso-Corrado et al., 2017). Este trabajo sugiere, por un lado, la alta sensibilidad y vulnerabilidad que presenta la especie a cambios en el ambiente y por otro, revela cómo la dinámica poblacional de la especie puede cambiar de un día a otro, debido a un evento extremo de fuertes vientos que azotó la región (Clark-Tapia et al., 2016). La ocurrencia de eventos extremos puede reducir drásticamente la abundancia de una especie, e incluso extinguirla localmente (Macedo-Ivanauskas, Monteiro y Ribeiro-Rodrigues, 2003). El estudio demográfico realizado en San Bernardo, durante el período 2011-2015 mostró una población en crecimiento de O. mexicana y sin amenaza o peligro de extinción desde una perspectiva demográfica, e incluso desde un enfoque genético de acuerdo con Naranjo-Luna (2014) y Pascual-Mendoza (2014).
Las poblaciones estudiadas durante este período en el bosque de niebla de Santiago Comaltepec, uno de los más conservados y grandes de México, mostraron que los fragmentos o parches analizados donde ocurre O. mexicana eran monodominandos por la especie, y sus tasas vitales permanecían estables y sin decremento poblacional aparente, al mostrar una estructura poblacional con regeneración favorable y un sistema maduro persistente (forma J-invertida) (Naranjo-Luna, 2014; Alfonso-Corrado et al., 2017). Al igual que para otras especies relicto (Hampe y Arroyo, 2002; Picó y Ribal, 2002; García, 2003), se encontró que los factores condicionantes de mortalidad disminuyen conforme las categorías de tamaño aumentan; patrón típico también de árboles de vida larga (Tlapa-Almonte, 2005; Freitas de Souza, Souza, Pizo, y Ganade, 2010). Una alta mortalidad en plántulas se atribuye principalmente a procesos densodependientes y a una selección de micrositios no favorables para el establecimiento (Pacheco-Cruz, 2016), mientras que en árboles adultos se asocia con enfermedades y derribos por fuertes vientos debido a que poseen un sistema radicular muy superficial en un suelo poco profundo (Naranjo-Luna, 2014).
No obstante que el análisis demográfico se realizó solamente en San Bernardo, la similitud estructural entre las diversas poblaciones locales permite vislumbrar un comportamiento de crecimiento estable para la especie, consistente con otras especies relicto cuyas poblaciones locales crecen en fragmentos (Hampe y Arroyo, 2002; Picó y Ribal, 2002; García, 2003). En San Bernardo, la variabilidad espacio temporal al valor de lambda se asocia predominantemente a la permanencia; similar a lo encontrado en especies relicto o endémicas, este parámetro demográfico, es fundamental para la persistencia de una especie (Clark-Tapia et al., 2006; García, 2008). Lo anterior se refleja en el análisis de elasticidad donde este parámetro demográfico es el más importante para O. mexicana y cuya dinámica poblacional depende de la supervivencia de los individuos adultos, en congruencia con lo publicado para especies de vida larga (Silvertown, Franco, Pisanty y Mendoza, 1993; Caswell, 2001) y para especies del orden fagales (Bonfil, 1998; Tlapa-Almonte, 2005; Alfonso-Corrado et al., 2007).
Por ser una especie eje y considerada una especie bandera del bosque de niebla en Santiago Comaltepec (Naranjo-Luna, 2014), se analizó el impacto de extracciones futuras de individuos adultos al crecimiento de la población. El uso de simulaciones en matrices de proyección en estudios demográficos permite establecer estrategias de conservación de una especie, al identificar los límites de extracción o pérdida que una población puede tolerar (Ticktin, Nantel, Ramírez y Johns, 2002). Las simulaciones de cosecha en San Bernardo muestran una alta vulnerabilidad de la especie en términos demográficos, cuando muere entre 25% y 50% de árboles adultos, al sufrir una reducción por debajo de la unidad de la tasa finita de crecimiento poblacional. Este resultado es similar al de otras especies de vida larga del orden Fagales, donde simulaciones de cosecha menores a 15% ocasionan un decremento drástico por debajo de la unidad en lambda, por lo que se recomienda un manejo local y extracciones mínimas (Alfonso-Corrado et al., 2007).
En 2016, el comportamiento demográfico O. mexicana cambio radicalmente debido a un evento climatológico de fuertes vientos (>100 km/h) ocurrido el 9 de marzo de 2016 (Clark-Tapia et al., 2016). En este evento, el tamaño de la población de diversos fragmentos, entre ellos San Bernardo, La Cañada y El Panteón decreció en más de 60% y murió por derribo más de 80% de los individuos adultos. En San Bernardo, la tasa finita de crecimiento poblacional disminuyó a 0.44 y pasó de tener una población en crecimiento en 2015 a una población en decremento.
El análisis demográfico del período 2011-2015 proyectó que la permanencia era el parámetro demográfico más importante para la persistencia local, con una alta vulnerabilidad a la muerte de individuos adultos, proyección que se confirmó en 2016. La permanencia de adultos en la categoría 3 representa en el análisis de elasticidad del período 2015-2016; nuevamente el parámetro demográfico más importante, congruente con la persistencia de especies amenazadas (Clark-Tapia et al., 2006; García, 2008). En este contexto, si se desea restaurar estas poblaciones locales, el esfuerzo de conservación debe enfocarse a lograr mantener y aumentar a futuro el número de individuos adultos.
Eventos extremos como el ocurrido en Santiago Comaltepec pueden ocasionar la disminución del tamaño poblacional a nivel local, o pueden originar la extinción local de una especie como lo menciona Macedo-Ivanauskas et al. (2003). A pesar de que el tamaño poblacional en diversos fragmentos de O. mexicana sufrió una drástica reducción en 2016, en este momento no se puede clasificar a la especie como en peligro de extinción a nivel local. Esto a pesar de que la especie sea especialista y necesite condiciones ambientales muy específicas (Naranjo-Luna, 2014; Alfonso-Corrado et al., 2017), o de que presente baja germinación y sus plántulas sean sensibles a cambios en la radiación dentro del bosque (Pacheco-Cruz, 2016). En este trabajo se sigue considerado una especie amenazada de acuerdo con el método de evaluación de riesgo de extinción de la Norma Oficial Mexicana NOM-059-SEMARNAT-2010; sin embargo, es necesario incorporar en futuras investigaciones el efecto de la variabilidad ambiental (e.g.Ticktin et al., 2002). Esto permitirá evaluar los factores que contribuyen a modificar el hábitat y a incrementar la fragmentación de las poblaciones estudiadas en subpoblaciones más pequeñas y aisladas, así como el posible impacto en la demografía de la especie.
Conclusiones
Este estudio concluye que la especie relicto y amenazada Oreomunnea mexicana, en la región de la Chinatla en Oaxaca, tiene una alta sensibilidad y vulnerabilidad a cambios ambientales extremos, que ocasionan cambios radicales en la estabilidad demográfica de la especie y amenazan la persistencia de las poblaciones locales. Para O. mexicana, la permanencia de los individuos es el parámetro demográfico con mayor contribución relativa al valor de λ y fundamental para la continuidad de las poblaciones locales de la especie. Al ser una especie de interés cultural y biológico las áreas donde se ubica se encuentran bajo protección comunitaria; por lo anterior, si se desea restaurar estas poblaciones, el esfuerzo de conservación debe enfocarse a lograr mantener y aumentar a futuro el número de individuos adultos en la zona.

