










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkHuitzil
versión On-line ISSN 1870-7459
Huitzil vol.9 no.2 Omitlán jul./dic. 2008
Resultados de monitoreos
Presencia y abundancia de aves de la isla Farallón de San Ignacio, Sinaloa
Presence and abundance of avian species on Farallón de San Ignacio Island, Sinaloa
Présence et abondance d'oiseaux sur l'île Farallón de San Ignacio, Sinaloa
Miguel Ángel Guevara-Medina1, José Alfredo Castillo-Guerrero1* y Marco Antonio González-Bernal2
1 Centro de Investigación Científica y de Educación Superior de Ensenada. Km. 107 Carretera Tijuana-Ensenada. A.P. 360. Ensenada, B.C. 22860, México. Correo electrónico: *acastill@cicese.mx.
2 Escuela de Biología. Universidad Autónoma de Sinaloa. Ciudad Universitaria, Culiacán, Sinaloa, México.
Recibido: 15 de julio de 2008.
Revisión aceptada: 2 de octubre de 2008.
Editor asociado: Horacio Jesús de la Cueva Salcedo.
Resumen
Para determinar la presencia y abundancia de aves que utilizan la isla Farallón de San Ignacio, Sinaloa, documentamos las aves observadas durante 46 visitas de cinco días a la isla entre 2003 y 2008. Registramos 57 especies de aves. En la isla anidaron regularmente cinco especies de aves marinas: Sula nebouxii, S. leucogaster, Phaethon aethereus, Phalacrocorax auritus y Larus heermanni. En 2008 anidó una rapaz, Falco peregrinus. Además, observamos 24 especies de aves acuáticas no reproductoras y 27 especies de aves terrestres. La temporada reproductiva de las especies anidantes coincide con la época de alta productividad primaria en el sur del Golfo de California. La isla es importante como sitio de anidación para P. aethereus y es una de las dos colonias más importantes en el Golfo de California. Para el resto de las especies anidantes, la isla tiene una importancia marginal, pues otras islas albergan colonias mayores. Debido a la ausencia de vegetación, las aves terrestres usan la isla sólo de manera ocasional y por breves periodos, especialmente los migrantes neotropicales durante la migración. Al comparar con otras islas de la parte sur del Golfo de California, Farallón de San Ignacio exhibió una riqueza de especies mayor a lo esperado de acuerdo con su tamaño. Esta discordancia se podría explicar por un esfuerzo mayor en el muestreo y posiblemente por la existencia de una mayor cantidad de especies de aves terrestres usando los hábitats costeros de Sinaloa y dispersándose ocasionalmente a islas cercanas.
Palabras clave: Islas, sur del Golfo de California, listado de aves, Phaethon aethereus, reproducción de aves marinas.
Abstract
To determine the presence and abundance of avian species that use Farallón de San Ignacio Island, Sinaloa, we documented the birds observed during 46 visits of five days on the island between 2003 and 2008. We registered 57 bird species. Five marine species regularly nested on the island: Sula nebouxii, S. leucogaster, Phaethon aethereus, Phalacrocorax auritus and Larus heermanni. In 2008, one raptor, Falco peregrinus, nested on the island. Added to which, we observed 24 species of non-nesting aquatic birds, and 27 species of terrestrial birds. The breeding season for nesting birds coincided with the period of high primary productivity in the Gulf of California. The island is an important nesting site for P. aethereus, and is one of the two most important colonies in the Gulf of California. The island is only marginally important for the rest of the nesting species as other islands have larger colonies. Due to the lack of vegetation, terrestrial birds use the island only occasionally and for brief periods, particularly neotropical migratory species during the migration. On comparing with other islands in the southern part of the Gulf of California, Farallón de San Ignacio exhibited greater species richness than expected for its size. This discordance could be explained by the greater sample effort, and the existence of a larger quantity of terrestrial bird species using coastal habitats in Sinaloa that may occasionally disperse to nearby islands.
Key words: Islands, southern Gulf of California, Bird list, Phaethon aethereus, reproduction of marine birds.
Resumé
Afin de déterminer la présence et l'abondance des oiseaux qui utilisent l'île Farallón de San Ignacio, Sinaloa, nous avons réalisé des observations au cours de 46 visites de cinq jours sur l'île, entre 2003 et 2008. Nous avons enregistré 57 espèces d'oiseaux. Sur l'île, cinq espèces d'oiseaux marins ont niché régulièrement : Sula nebouxii, S. leucogaster, Phaethon aethereus, Phalacrocorax auritus et Larus heermanni. En 2008, une espèce de rapace, Falco peregrinus, a niché. De plus, nous avons observé 24 espèces d'oiseaux aquatiques non reproductrices et 27 espèces d'oiseaux terrestres. La saison reproductive des espèces qui ont niché coïncide avec la saison de production primaire élevée dans le sud du Golfe de Californie. L'île est un important site de nidification pour P. aethereus et abrite l'une des plus grosses colonies du Golfe. Pour le reste des espèces nicheuses, l'île revêt une importance marginale puisque d'autres îles abritent des colonies plus grandes. À cause de l'absence de végétation, les oiseaux terrestres utilisent l'île de manière sporadique seulement, notamment les espèces néotropicales en migration. Comparée à d'autres îles du sud du Golfe de Californie, Farallón de San Ignacio exhibe une richesse spécifique plus élevée en relation avec sa superficie, qui pourrait s'expliquer par l'effort d'échantillonnage plus grand et possiblement par la présence plus nombreuse d'espèces d'oiseaux terrestres qui utilisent les habitats côtiers du Sinaloa et se dispersent occasionnellement sur les îles proches.
Mots clé: Îles, sud du Golfe de Californie, listing des oiseaux, Phaethon aethereus, reproduction d'oiseaux marins.
Introducción
Aunque las islas del Golfo de California están consideradas como áreas naturales protegidas desde 1978 (D.O.F. 1978), la información básica de sus recursos bióticos es limitada. En el caso de las aves existen algunos estudios que han documentando los patrones de ocurrencia y abundancia en algunas islas y se ha tratado de entender sus afinidades biogeográficas (Cody y Velarde 2002), pero el esfuerzo ha sido relativamente limitado pues la cantidad de islas (aproximadamente 900 islas e islotes) rebasa las capacidades de los grupos académicos en la región. Así, el conocimiento básico de las aves que habitan las islas en el Golfo de California es pobre y en muchos casos, las listas de especies están aún incompletas (Cody y Velarde 2002).
Para la porción sur del Golfo de California, los estudios se han concentrado en islas cercanas a la península de Baja California como Cerralvo (Banks 1963), Carmen (Gaviño et al. 1984, Cody y Velarde 2002) y Espíritu Santo (Carmona et al. 2005), por lo que la carencia de información es aún más acentuada para las islas cercanas a la costa de Sinaloa.
En el noroeste de México, durante las ultimas décadas, ha aumentado el acceso y atractivo de las islas para pescadores comerciales, turistas y otros visitantes, por lo que el número de gente usando las islas y los problemas asociados con el uso humano parecen haberse incrementado (Velarde y Anderson 1994, Tershy et al. 1997). Por tanto, el generar datos que permitan entender los patrones de uso de los diferentes grupos zoológicos en las islas, permitirá una mejor planificación de su uso y la implementación de acciones de conservación efectivas, así como evaluar con mayor certidumbre los efectos del uso humano en sitios que hasta recientemente se consideraban como "uno de los archipiélagos no polares, ecológicamente más intactos" (Tershy y Breese 1997).
En este sentido, presentamos una lista de las especies observadas entre 2003 y 2008 en la isla Farallón de San Ignacio, en el sureste del Gofo de California. Compilamos y complementamos la información generada previamente, basada en visitas cortas (Everett y Anderson 1991, González-Bernal et al. 2002, Mellink et al. 2002), incluimos comentarios sobre las temporadas de anidación, número de individuos observados, éxito reproductivo y dieta de las especies anidantes, y frecuencia de observación y abundancia de las especies no reproductoras. Además, realizamos una comparación con información publicada sobre otras islas en el sur del Golfo de California.
Métodos
Área de estudio
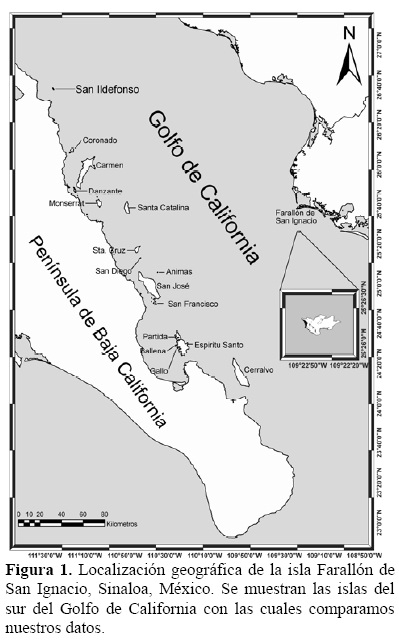
Farallón de San Ignacio (25° 26' N, 109° 22' W; hasta 140 msnm) es un pequeño islote desprovisto de vegetación, localizado en la parte suroriental del Golfo de California, a 36 km de la costa de Topolobampo, en el norte de Sinaloa (Figura 1). Tiene un área aproximada de 16 ha (Samaniego-Herrera et al. 2008). En la cima hay una planicie de unas 3.5-4 ha (González-Bernal et al. 2002) rodeada por acantilados (Figura 2). La isla se encuentra cercana a la cuenca Farallón y está rodeada de aguas profundas (sensu Maluf 1983). La temperatura media anual es superior a los 22°C, con una oscilación térmica superior a los 14°C y temperaturas superiores a los 18°C en invierno. La precipitación pluvial es de 310.5 mm con un periodo de lluvias en verano, principalmente en el mes de agosto (Gutiérrez 2003 en Peñaloza 2006).
Trabajo de campo
Realizamos dos visitas mensuales de cinco días cada una, entre enero y mayo de 2003, 2004, 2006 y 2007; además de visitas adicionales en octubre de 2004, marzo de 2005 y junio de 2006. En 2008 visitamos la isla una vez por mes entre marzo y mayo. Durante cada visita se tomó nota de las aves observadas. En el caso de las aves anidantes se calculó el número de adultos y en el caso de Phaethon aethereus, Sula nebouxii y S. leucogaster, se determinó el éxito reproductivo monitoreando nidos a lo largo de la temporada. Se tomaron notas sobre las presas consumidas por cada especie observando y colectando regurgitados.
Para comparar el número de especies observadas con otras islas de la porción sur del Golfo de California usamos la información publicada por Cody y Velarde (2002), complementada con información más reciente sobre algunas islas y con aquella publicada sobre especies reproduciéndose (Carmona et al. 1994, Carmona et al. 1996, Patten 1996, Mellink 2001, Mellink et al. 2002, Carmona et al. 2005, Anderson et al. 2007, Mellink et al. 2007). El área y la distancia de cada isla con respecto a la península o al continente fueron tomadas de Murphy et al. (2002) y para las islas en que no encontramos datos, las calculamos usando el programa Google Earth (versión 4.2.0180.1134).
El orden y la nomenclatura en que presentamos el listado es el sugerido por la AOU (1998), considerando las actualizaciones publicadas como suplementos en la revista Auk (42-49).
Resultados
A continuación enlistamos las aves observadas y se agregan comentarios sobre las especies que anidan y sobre aquellas cuyo registro es notable a escala regional.
Especies reproductoras
Bobo patazul (Sula nebouxii)
Es el ave anidante más abundante en la isla. Su temporada reproductiva abarca de diciembre a mayo y en ocasiones se alarga hasta junio; la puesta de huevos ocurre regularmente en enero y el periodo de eclosiones a fines de febrero. El número de parejas fluctúa entre 1,500 y 2,500. La mayoría de los nidos se colocan en la cima de la isla alcanzando densidades de hasta 0.6 nidos m-2, aunque hay nidos dispersos en todos los acantilados de la isla. El éxito reproductivo osciló entre el abandono de todos los nidos en 2003 y un promedio ligeramente mayor a un volantón por nido (1.17 volantones nido-1) en 2006. Su dieta la componen principalmente peces pelágicos menores, los más importantes son la anchoveta (Cetengraulis mysticetus) y la sardina crinuda (Ophistonema sp.) con una frecuencia de aparición en los regurgitados del 55% y 10% respectivamente (n=80, durante la temporada 2004)
Bobo café (Sula leucogaster)
Es un anidante abundante presente todo el año. Su temporada reproductiva abarca de noviembre a abril o mayo. Anidan aproximadamente 1,200 parejas (González-Bernal et al. 2002) que usan principalmente los acantilados o zonas con inclinación pronunciada; aunque hay algunos nidos en la cima mezclados con S. nebouxii. El hecho de anidar en la cima, donde hay una alta densidad de S. nebouxii, puede ocasionar que ocurra hibridación entre estas especies; incluso, se han reportado aves con caracteres intermedios en la isla (Castillo-Guerrero et al. 2005). Las presas principales de S. leucogaster en Farallón de San Ignacio son las anchovetas (C. mysticetus) y cornetas (Fistularia corneta); aunque su dieta es muy variada y puede incluir sardinas (Ophistonema sp.), macarelas (Scomber japonicus), peces voladores (Decapterus sp.) y pajaritos (Hemiramphus sp., Peñaloza 2006).
Rabijunco pico-rojo (Phaethon aethereus)
Es un anidante común en la isla. Previamente se calcularon entre 150 y 200 parejas (González-Bernal et al. 2002). Observamos que hay variaciones interanuales importantes en la ocupación de nidos, aunque el total que calculamos fue similar al de González-Bernal et al. (2002), con un máximo de 250 a 300 parejas en 2008. La temporada de anidación ocurre entre octubre y mayo, presentando dos picos de puesta durante los meses de noviembre y febrero. Cada nido contiene un sólo huevo y el éxito reproductivo es variable entre temporadas (entre 0.49 volantones nido-1 en 2007 y 0.95 volantones nido-1 en 2008). Para anidar, utiliza las cavidades que hay entre las rocas en todas las caras de la isla, excepto en la cima. Las especies mas comunes en su dieta son el pez volador (Fodiator acutus), calamar (Loliolopsis diomedeae) y langostilla (Pleuroncodes planipes). Antes de nuestro estudio se creía que la isla San Pedro Mártir albergaba la colonia más importante para la especie en el Golfo de California (Everett y Anderson 1991, Tershy y Breese 1997); sin embargo, la cantidad de parejas en Farallón de San Ignacio es de una magnitud similar y en conjunto, las dos islas contienen al menos la mitad de aves calculadas por Everett y Anderson (1991) para el Golfo de California.
Cormorán orejudo (Phalacrocorax auritus)
Es un anidante común. Esta especie construye alrededor de 60-90 nidos por año. Anida de octubre a Marzo en el cantil norte de la isla. Debido a su susceptibilidad al disturbio, no se colectó información sobre dieta y éxito reproductivo de esta especie.
Gaviota ploma (Larus heermanni)
Es un anidante común, presente todo el año. Calculamos alrededor de 2,000 parejas anidando entre marzo y mayo. La mayoría anidan en la cara oeste de la isla, aunque se observan nidos en la cima y en los cantiles del resto de la isla. Una presa común en su dieta es la langostilla (Pleuroncodes planipes); también se alimenta cleptoparasitando a S. nebouxii y S. leucogaster, especialmente cuando los adultos intentan alimentar a pollos con más de cinco semanas de edad.
Halcón peregrino (Falco peregrinus)
Anidante raro en la isla. Entre 2003 y 2007 realizamos varias observaciones de individuos perchando en los acantilados y volando sobre la isla. En marzo de 2008 observamos una pareja en el acantilado oeste exhibiendo comportamiento de defensa de nido. En abril de 2008 localizamos el nido, el cual contenía dos huevos. Aparentemente, 2008 fue el primer año registrado de anidación de la especie en la isla. La anidación de esta especie se ha registrado previamente en varias islas del Golfo de California como San Pedro Martír, Espíritu Santo y Rasa (Banks 1969, Tershy y Breese 1997).
Registros notables
Bobo pata-roja (Sula sula)
Previamente reportado para la isla en González-Bernal et al. (2007). Observamos un individuo descansando en la cima de la isla, junto a un grupo de juveniles de S. nebouxii el 24 de octubre de 2004. Sólo había un registro previo dentro del Golfo de California, de manera que fue el primer reporte para Sinaloa.
Bobo de Nazca (Sula granti)
El 9 de mayo de 2006, observamos un individuo con plumaje intermedio entre juvenil y adulto, el cual lucía gastado, lo que indicó que la muda al plumaje adulto podría estar próxima. El pico era amarillo, aunque de un tono naranja en la base, las patas grises, el inicio de la máscara aún pálida y grisácea, el manto claro con plumas cafés intercaladas, cabeza predominantemente blanca con las puntas de las plumas cafés y la pupila amarilla; las retrices centrales eran blancas excepto en la base. Se encontraba descansando con un grupo de volantones de S. nebouxii, el tamaño era claramente mayor (≈ 1.3 veces). El primero de junio de 2008 observamos otro individuo subadulto al cual ya se le notaba el color del pico naranja rosado, que lo diferencia del S. dactylatra que tiene el pico amarillo. Había dos registros previos en el Golfo de California, uno en la Isla San Pedro Mártir (Tershy y Breese 1997) y otro cerca de Mazatlán (Huyvert y Anderson 2004). Nuestros registros confirman la presencia ocasional de la especie dentro del Golfo de California, principalmente de individuos inmaduros.
Gaviota patamarilla (Larus livens)
Es una especie común todo el año en la isla. Aunque previamente se consideró que su reproducción era probable (González-Bernal et al. 2002), nosotros no observamos nidos durante nuestras visitas. Sin embargo, en abril de 2008, observamos una pareja acarreando material y acomodándolo para formar la estructura del nido. Es común ver a esta especie depredando huevos de S. nebouxii, S. leucogaster y L. heermanni, o cleptoparasitando a las mismas especies. Esta especie es endémica del Golfo de California, sus colonias en la porción sur del Golfo se ubican exclusivamente en islas cercanas a la península de Baja California (Patten 1996), por lo que de confirmar su anidación sería el primer registro en Sinaloa.
Las especies sin reproducción observada se muestran en el cuadro 1.
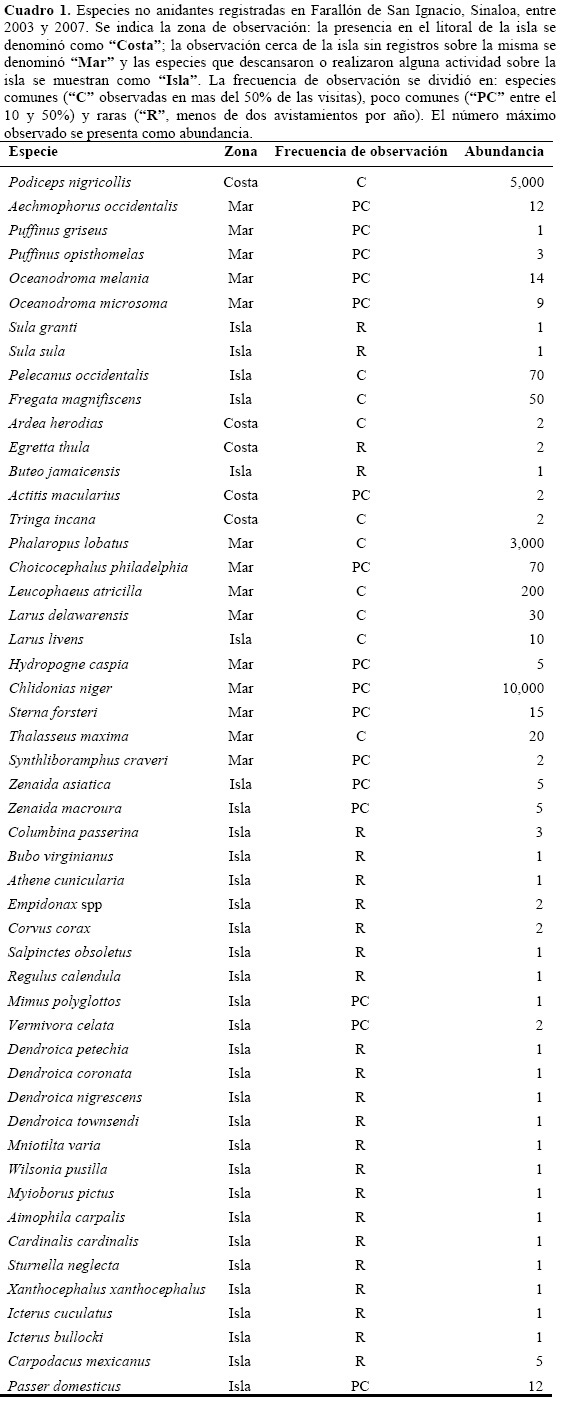
Al comparar con otras islas del sur del Golfo de California (Cuadro 2), se observa que las 57 especies observadas en el Farallón de San Ignacio equivalen a las observadas en islas con extensiones geográficas considerablemente mayores. Esta diferencia resalta especialmente en los grupos de aves terrestres no reproductoras y de marinas no reproductoras. Farallón de San Ignacio junto con San Ildefonso son las islas con más especies de aves marinas anidantes en la sección considerada del sur del Golfo de California (Cuadro 2).
Discusión
La temporada reproductiva de las aves marinas en la isla esta fuertemente influenciada por las condiciones oceanográficas de la zona. Todas las especies anidan durante la época de alta productividad primaria en el sur del Golfo de California. Esta ocurre de noviembre a mayo, cuando el viento ocasiona surgencias en el margen este del golfo, lo que incrementa la productividad general en la región (Álvarez-Borrego 2002). En aves como P. aethereus, el inicio de la puesta de huevos coincide con un decremento, a menos de 27 °C, en la temperatura superficial del mar en los alrededores de la isla (Guevara 2008). Además, las variaciones interanuales en las condiciones oceanográficas tienen efectos en el desempeño reproductivo de las especies anidantes; por mencionar un ejemplo, el éxito reproductivo de S. nebouxii en la isla disminuye cuando el promedio anual de la temperatura superficial del mar aumenta (J.A.C.-G. datos no publicados).
El hecho de que la isla se encuentre cercana a la costa este del Golfo de California, donde se originan las surgencias en invierno y donde ocurre la mayor productividad primaria en la porción sur del golfo, podría explicar, al menos parcialmente, que a escala regional sea una de las dos islas con mayor cantidad de especies anidando. Otra condición favorable que se cumple en las dos islas con mayor cantidad de aves marinas anidantes (Farallón de San Ignacio y San Ildefonso), es la ausencia de mamíferos nativos depredadores (como Basariscus astutus en Espíritu Santo y San José o Canis latrans en San José) y de gatos introducidos (como en el resto de las islas consideradas en la comparación (sensu Lawlor et al. 2002).
Las 27 especies de aves terrestres que observamos en la isla la usan de manera ocasional, especialmente de febrero a abril, lo cual coincide con la época de migración hacia las zonas de reproducción en el norte. Aparentemente, la isla es usada sólo como zona de descanso durante la migración. En el Golfo de California hay pocas islas a mayor distancia de la península o del continente que Farallón de San Ignacio (Cuadro 2), de manera que el observar aves terrestres en esta isla pequeña y sin hábitat adecuado para su permanencia pone de manifiesto sus capacidades de dispersión; sin embargo, el tiempo de estancia dependerá entonces de los requerimientos de hábitat de cada especie y su disponibilidad en las diferentes islas.
El total de especies observadas en Farallón de San Ignacio es discordante con el patrón observado en las islas del margen oeste del golfo, donde la riqueza de especies se correlaciona con el tamaño de la isla (Cody y Velarde 2002, Cuadro 2). Las 57 especies que nosotros reportamos están muy por encima de lo esperado de acuerdo con el tamaño de la isla; esto es especialmente notable al considerar que la isla está desprovista de vegetación y que está relativamente alejada de la costa. El hecho de tener una riqueza comparable con la de islas con extensiones mayores en dos órdenes de magnitud puede tener dos causas no excluyentes:
1) Que sea producto de un esfuerzo de muestreo mayor al realizado en las otras islas. Esto parece cierto para las aves marinas no reproductoras, pues no encontramos listados con información sobre este grupo para ninguna de las islas cercanas a la península de Baja California, pese a que muchas de las especies que nosotros observamos en Farallón de San Ignacio son de distribución amplia en la región (e.g. Larinae y Sterninae).
2) Por otra parte, las islas de las que se tiene información se encuentran cercanas a la península de Baja California, donde predomina una vegetación de desierto mientras que en el lado este, en el norte de Sinaloa, la precipitación es mayor a los 300 mm y la vegetación se torna más densa, alta y arbustiva, es decir, hay una transición de desierto a bosque espinoso (Johnson 1982, Cody y Velarde 2002). Esto tiene como consecuencia una mayor productividad general (Cody y Velarde 2002), por lo que hay mayor cantidad de recursos disponibles para las aves terrestres. Así, aunque en la isla no se cumple estrictamente lo anterior, pues no hay vegetación, el uso de la zona costera de Sinaloa por una mayor cantidad de especies aumentaría la probabilidad de que éstas eventualmente alcancen las islas más cercanas. Aunque no hay información de otras islas en el margen este del golfo para corroborar este patrón, la riqueza de aves, particularmente terrestres, parece ser mayor en las islas del margen este del Golfo de California que en islas cercanas a la península de Baja California.
Durante nuestras visitas, un problema de conservación evidente fue la presencia de ratas (Rattus rattus). Aparentemente fueron introducidas en el siglo pasado, durante la explotación de guano en la isla (Gonzalez-Bernal et al. 2002) y se cree que pudieron modificar la comunidad de especies anidantes excluyendo por depredación a aves de tamaño pequeño como petreles, pardelas y mergulos (sensu McChesney y Tershy 1998, Donlan et al. 2003). El efecto de las ratas en la isla es una especulación, pues no se cuenta con datos previos a la introducción; sin embargo, se sabe que afectan negativamente a las aves marinas (al menos en 61 islas en todo el mundo) y se cree que son responsables de numerosas extirpaciones y decrementos poblacionales (Jones et al. 2008). Como medida de restauración ecológica, en el otoño de 2007, el Grupo de Ecología y Conservación de Islas (GECI) aplicó un protocolo para la erradicación de ratas (R. rattus) en esta isla (Samaniego-Herrera et al. 2008). Al abarcar los años previos a la erradicación con un esfuerzo considerable, este trabajo puede servir como punto de comparación para monitorear posibles cambios en la estructura de la comunidad de aves anidantes y ayudar a determinar los efectos de la erradicación a diferentes escalas temporales.
Agradecimientos
A CONACyT (SEP-2004-C01-45995), CICESE y GECI, por su apoyo financiero para realizar las salidas de campo. A E. Peñaloza, M. Prado y F. Cárdenas por su apoyo en el trabajo de campo. A E. Mellink y las familias González-Bernal y Aguilar-Camacho por su apoyo logístico. A E. Mellink, H. de la Cueva y un revisor anónimo por sus comentarios y su ayuda en la edición del manuscrito.
Literatura citada
AOU. (American Ornithologists' Union). 1998. Checklist of North American Birds, 7a ed. American Ornithologists Union. Washington, D.C. [ Links ]
Alvarez-Borrego, S. 2002. Physical oceanography. Pp. 41-59. In: T.J. Case, M.L. Cody y E. Ezcurra (eds.). A new Island Biogeography of the Sea of Cortés. Oxford University Press. Nueva York. [ Links ]
Anderson, D.W., C.J. Henny, C. Godinez-Reyes, F. Gress, E. Palacios, K. Santos del Prado y J. Bredy. 2007. Size of the California brown pelican metapopulation during a Non-El Niño Year: Geological Survey, Open-File Report 2007-1299. Reston, Virginia, E.U.A. [ Links ]
Banks, R.C. 1963. Birds of Cerralvo Island, Baja California. Condor 65:300-312. [ Links ]
Banks, R.C. 1969. The Peregrine Falcon in Baja Caifornia and the Gulf of California Pp. 81-91. In: J.J. Hickley (ed.). Peregrine Falcon populations: their biology and decline. University of Wisconsin Press. Madison. [ Links ]
Carmona, R., J. Guzman, S. Ramirez y J. Fernandez. 1994. Breeding waterbirds of La Paz Bay, Baja California Sur, México. Western Birds 25:151-157. [ Links ]
Carmona, R., S. Ramirez, B. Zarate y F. Becerril. 1996. Some nesting waterbirds from southern San José island and adjacent islands, Gulf of California, México. Western Birds 27:81-85. [ Links ]
Carmona, R., G. Ruiz-Campos, J.A. Castillo-Guerrero y G. Brabata. 2005. Patterns of occurrence and abundance of landbirds on Espíritu Santo Island, Gulf of California, México. Southwestern Naturalist 50:440-447. [ Links ]
Castillo-Guerrero, J.A., E. Mellink, E.A. Peñaloza-Padilla y M. Prado-López. 2005. Anomalously pigmented Brown Boobies in the Gulf of California: Leucism and possibly hybridization with the Blue-footed Booby. Western Birds 36:325-328. [ Links ]
Cody, M.L. y E. Velarde. 2002. Land birds. Pp. 271-312. In: T.J. Case, M.L. Cody y E. Ezcurra (eds.). A new Island Biogeography of the Sea of Cortés. Oxford University Press. Nueva York. [ Links ]
D.O.F. (Diario Oficial de la Federación). 1978. Decreto por el que se establece una zona de reserva y refugio de aves migratorias y de la fauna silvestre, en las islas que se relacionan, situadas en el Golfo de California. 2 de agosto de 1978. México, D. F. [ Links ]
Donlan, C.J., G.R. Howald, B. Tershy y D.A. Croll. 2003. Evaluating alternative rodenticides for island conservation: roof rat eradication from the San Jorge Islands, Mexico. Biological Conservation 114:29-34. [ Links ]
Everett, W.T. y D.W. Anderson. 1991. Status and conservation of the breeding seabirds on offshore pacific islands of Baja California and the Gulf of California. International Council of Bird Protection Tecnical Publication 11:115-139. Cambridge. [ Links ]
Gaviño, G., F. Aguilar y L.C. García. 1984. Abundancia relativa, reproducción y actividad diurna de aves terrestres en la Isla Carmen, Baja California Sur, México. Anales del Instituto de Biología, Universidad Nacional Autónoma de México, Serie Zoología 55:263-284. [ Links ]
González-Bernal, M.A., E. Mellink y J.R. Fong-Mendoza. 2002. Nesting birds of Farallón de San Ignacio, Sinaloa, Mexico. Western Birds 33:254-257. [ Links ]
González-Bernal, M.A., J.A. Castillo-Guerrero, C.R. Hernández-Celis y E. Mellink. 2007. Noteworthy bird records from Sinaloa, Mexico. Western Birds 38:52-56. [ Links ]
Guevara, M.A. 2008. Variación interanual en los patrones reproductivos del rabijunco de pico rojo (Phaethon aethereus) en las temporadas 2004 y 2007 en la isla Farallón de San Ignacio, Sinaloa. Tesis de maestría, Centro de Investigación Científica y de Educación Superior de Ensenada. Ensenada, Baja California. [ Links ]
Huyvaert, K.P. y D.J. Anderson. 2004. Limited dispersal by Nazca Boobies Sula grantii. Journal of Avian Biology 35:46-53. [ Links ]
Johnson, A.F. 1982. Dune vegetation along the eastern shore of the Gulf of California. Journal of Biogeography 9:317-330. [ Links ]
Jones, H.P., B.R. Tershy, E.S. Zavaleta, D.A. Croll, B.S. Keitt, M.E. Finkelstein y G.R. Howald. 2008. Severity of the effects of invasive rats on seabirds: a global review. Conservation Biology 22:16-26. [ Links ]
Lawlor, T.E., D.J. Hafner, P.T. Stapp, B.R. Riddle y S.T. Alvarez-Castañeda. 2002. Mammals. Pp. 326-361. In: T.J. Case, M.L. Cody y E. Ezcurra (eds.). A new Island Biogeography of the Sea of Cortés. Oxford University Press. Nueva York. [ Links ]
Lee, D.S. y M. Walsh-Mcgehee. 2000. Populations estimates, conservation concerns, and management of tropicbirds in the western Atlantic. Caribbean Journal of Science 36:267-279. [ Links ]
Maluf, L.Y. 1983. Physical oceanography. Pp. 26-45. In: T.J. Case y M.L. Cody (eds.). Island biogeography in the Sea of Cortez. University of California Press. Berkeley. [ Links ]
McChesney, G.J. y B. Tershy, B. 1998. History and status of introduced mammals and impacts to breeding seabirds on the California Channel and northwestern Baja California Islands. Colonial Waterbirds 21:335-347. [ Links ]
Mellink, E. 2001. History and status of colonies of Heermann's Gull in Mexico. Waterbirds 24:188-194. [ Links ]
Mellink, E., A. Orozco-Meyer, B. Contreras y M. González-Jaramillo. 2002. Observations on nesting seabirds and insular rodents in the middle Sea of Cortés in 1999 and 2000. Bulletin of the Southern California Academy of Sciences 101:28-35. [ Links ]
Mellink, E., E. Palacios y E. Amador. 2007. Colonies of four species of terns and the black skimmer in Western Mexico. Waterbirds 30:358-366. [ Links ]
Murphy, R.W., F. Sanchez-Piñedo, G.A. Polis y R.L. Aalbu. 2002. New measurements of area and distance for islands in the Sea of Cortés. Pp. 447-464. In: T.J. Case, M.L. Cody y E. Ezcurra (eds.). A new Island Biogeography of the Sea of Cortés. Oxford University Press. Nueva York. [ Links ]
Patten, M.A. 1996. Yellow-footed gull (Larus livens). The Birds of North America. (A. Poole, Ed.). Ithaca: Cornell Lab of Ornithology <http://bna.birds.cornell.edu/bna/species/243doi:10.2173/bna.243> (consultado 1 junio de 2008). [ Links ]
Peñaloza, E.A. 2006. Cuidado parental y ecología de forrajeo del bobo café (Sula leucogaster brewsteril) en el Farallón de San Ignacio, Sinaloa, México, en la temporada reproductiva 2004. Tesis de licenciatura, Universidad de Guadalajara. Guadalajara, Jalisco. [ Links ]
Samaniego-Herrera, A., A. Aguirre-Muñoz, M. Felix-Lizarraga, J. Valdez-Villavicencio, R. González-Gómez, M. Rodriguez-Malagon, A. Peralta-García, F. Torres-García, F. Mendez, N. Silva, E. Sodi, G. Howald y B. Tershy. 2008. Restoring island ecosystems in the Gulf of California: black rat erradication on Farallon de San Ignacio and San Pedro Martir Islands. Seventh California Islands Symposium. Embassy Suites. Oxnard, CA. [ Links ]
Tershy, B.R. y D. Breese. 1997. The birds of San Pedro Martir Island, Gulf of California, Mexico. Western Birds 28:96-107. [ Links ]
Tershy, B.R., D. Breese y D. Croll. 1997. Human perturbations and conservation strategies for San Pedro Mártir Island, Islas del Golfo de California Reserve, México. Environmental Conservation 24:261-270. [ Links ]
Velarde, E. y D.W. Anderson. 1994. Conservation and management of seabird islands in the Gulf of California: setbacks and successes. Pp 229-243. In: D.N. Nettleship, J. Burger y M. Gochfeld (eds.). Seabirds on islands: treats, case studies and action plans. ICPB technical publication. Cambridge. [ Links ]

